Сравниваем поведение автомобиля на колесах разных размеров — журнал За рулем
Как меняется поведение автомобиля в зависимости от размера шин? Чтобы ответить на этот вопрос, тестовая группа журнала «За рулем» провела серию замеров на автомобиле Volkswagen Golf, используя шины одной модели, но разного диаметра – 16 и 17 дюймов.
Допустим, на машине штатно установлены 17‑дюймовые покрышки, но автопроизводитель допускает и использование шин с посадочным диаметром на дюйм меньше. Отлично! Ведь более узкая шина меньшего диаметра дешевле порою на 25–30%! Даже с учетом покупки недорогих «зимних» дисков — есть смысл, к тому же потом на шиномонтаже сэкономить можно. Но не растеряют ли шины часть своих свойств при переходе на меньший диаметр?
Материалы по теме
На полигоне мы испытывали нешипованные шины Nokian Hakkapeliitta R2 двух размерностей — 205/55 R16 и 225/45 R17. Прогнали их по стандартной методике, по которой испытываем все зимние шины в сравнительных тестах. Дополнительно лишь проверили, на какой скорости начинается аквапланирование.
Дополнительно лишь проверили, на какой скорости начинается аквапланирование.
Без точных измерительных приборов поймать разницу в поведении автомобиля на «шестнадцатых» и «семнадцатых» колесах было бы крайне сложно. В большинстве случаев показатели различаются не более чем на пару процентов. А такой разброс вполне можно отнести на счет погрешности измерений. Более существенная разница проявляется при торможении на льду: покрышки R16 немного проигрывают «семнадцатым». Зато на «шестнадцатых» значительно выше скорость начала аквапланирования. По личному опыту могу сказать, что примерно такая же разница будет и на снежной каше.
В целом, полученные результаты замеров позволяют назвать сравниваемые шины равноценными. А что скажут эксперты? По оценке экспертов шинной группы «За рулем», почти во всех дисциплинах «маленькие» шины оказались лучше «больших». Например, очевидна разница в комфорте. Более узкие шины меньше шумят и мягче принимают дорожные неровности.
 zr.ru/d/story/77/905079/251.svg”/>
zr.ru/d/story/77/905079/251.svg”/>
При движении с высокой скоростью по заснеженной дороге руль на автомобиле, обутом в шины R16, немного плотнее, «ноль» более отчетливый. Разница в управляемости тоже минимальна, но опять-таки в пользу «шестнадцатых». На шинах R16 баранка чуть более информативна, поворачивать ее приходится на меньшие углы, поэтому Golf легче и четче управляется в скольжениях.
Материалы по теме
При оценке проходимости перевес вновь на стороне более узких шин — машина режет сугробы увереннее. А на широких она сталкивается с повышенным сопротивлением. Правда, среднестатистический водитель такую разницу вряд ли поймает. Хотя, скорее всего, почувствует, что на «шестнадцатых» управлять автомобилем на заснеженных дорогах надежнее и проще. А значит — безопаснее.
Вывод очевиден: шины 205/55 R16 лучше подходят для зимы, чем аналогичные размерностью 225/45 R17. Причем такой вывод сделают абсолютно все водители, независимо от уровня их подготовки. Главное — «шестнадцатые» обеспечат более надежное движение по снежной каше и лужам, а желающему прохватить с ветерком по заснеженным дорогам доставят больше удовольствия от управления автомобилем. Кроме того, «маленькие» шины дешевле и благодаря более высокому (читай: энергоемкому) профилю живут на наших неровных дорогах дольше.
Главное — «шестнадцатые» обеспечат более надежное движение по снежной каше и лужам, а желающему прохватить с ветерком по заснеженным дорогам доставят больше удовольствия от управления автомобилем. Кроме того, «маленькие» шины дешевле и благодаря более высокому (читай: энергоемкому) профилю живут на наших неровных дорогах дольше.
Шинный калькулятор
Подбор шин с помощью шинного калькулятора позволяет выбрать оптимальную модель покрышки. Шинный калькулятор служит для вычисления взаимозаменяемых размеров шин или, наоборот, проверки технической возможности установки на автомобиль шин не заводской размерности. Важнейшим параметром типоразмера является полный внешний диаметр колеса. При этом сама величина наружного диаметра покрышки не указана в европейском написании типоразмера шины и требует математического вычисления. В процессе подбора шин с помощью шинного калькулятора важно учитывать, что допустимо безболезненное отклонение полного диаметра покрышки до 2% от номинала и до 3% с обязательной примеркой изделий на дисках в сборе.
Не менее важным является ширина покрышки. Однако при подборе шин ее нельзя вычислить с помощью шинного калькулятора — только с помощью примерки. Можно только примерно предполагать среднее 10% отклонение. Хотя на внедорожниках большие колесные арки предполагают и куда более значительные отклонения. Лучше всего позвоните нам и получите подробную консультацию у опытного специалиста, чем осуществлять самостоятельный подбор шин. Нами почти все варианты для всех автомобилей уже давно примерены и опробованы.
Шинный калькулятор
Ширина:Все155165175185195205215225235245255265275285295305315325335345355385425435056.46.577.58.599.51010.51111.51212.51313.5212730313233355560708090100110120130135140145150160170180190200210220240260280300400500
Все253035404550556065707580859008. 59.51010.51111.51212.5313233100
59.51010.51111.51212.5313233100
Все13C131414C15C1516C161717.517C1818C1919.519C2020C212222.522.5C23185225012C122436533
Ширина:Все155165175185195205215225235245255265275285295305315325335345355385425435056.46.577.58.599.51010.51111.51212.51313.5212730313233355560708090100110120130135140145150160170180190200210220240260280300400500
Профиль:Все253035404550556065707580859008.59.51010.51111.51212.5313233100
Диаметр: Все13C131414C15C1516C161717. 517C1818C1919.519C2020C212222.522.5C23185225012C122436533
517C1818C1919.519C2020C212222.522.5C23185225012C122436533
| Результаты | Старый | Новый | Разница |
|---|---|---|---|
| L — ширина шины, мм | {{ form.width }} | {{ form.c_width }} | {{ form.c_width-form.width }} |
| H — высота профиля, мм | {{ height_mm=form.width*form.height/100 | number : 0 }} | {{ c_height_mm-height_mm | number : 0 }} | |
| d — внутренний диаметр, мм | {{ diametr_mm=25.4*form.diametr | number : 0 }} | {{ c_diametr_mm=25. 4*form.c_diametr | number : 0 }} 4*form.c_diametr | number : 0 }} | {{ c_diametr_mm-diametr_mm | number : 0 }} |
| D — внешний диаметр, мм | {{ full_diametr_mm = diametr_mm + height_mm * 2 | number : 0 }} | {{ c_full_diametr_mm = c_diametr_mm + c_height_mm * 2 | number : 0 | number : 0 }} | {{ delta_full_diametr_mm = c_full_diametr_mm – full_diametr_mm | number : 0 }} |
| Изменение клиренса, мм | {{ delta_full_diametr_mm / 2 | number : 0 }} | ||
| Разница в показаниях спидометра | {{ delta_full_diametr_mm / full_diametr_mm * 100 | number : 0 }} % | ||
И отталкиваться при изменении ширины покрышки при введении данных в шинный калькулятор желательно от среднего значения. Например, выбирая шины для внедорожников на большинстве элитных кроссоверов, таких как BMW X5, Porsche Cayenne, Volkswagen Touareg, Audi Q7, Subaru Tribeca, Volvo XC90, Range Rover Sport и т. п. средним является размер 255/55 R18, а заводскими взаимозаменяемыми могут быть как меньший посадочный!!! размер 235/65 R17 и 255/60 R17 так и эквивалентный, но более узкий 235/60 R18, так и 255/50 R19, 285/45 R19 в 19м посадочном диаметре, 245/45 R20, 275/40 R20 и даже 315/35 R20, и даже 22 дюймовые аналоги 285/30 R22. Так и для большинства легковых авто среднего класса суперпопулярный размер 205/55 R16 заменяем размерами 195/65 R15 и 205/60 R15 при уменьшении посадочного диаметра шины, 195/60 R16 и 225/50 R16 при необходимости изменения ширины покрышки, 205/50 R17 и 225/45 R17 для понижения профиля шин, 225/40 R18 и 255/35 R18 для еще более спортивного поведения автомобиля и 225/35 R19 и соответственно 255/30 R19 для самых экстремалов. Более широкие варианты покрышек предназначены для подбора шины для заднеприводных автомобилей Мерседес и БМВ.
п. средним является размер 255/55 R18, а заводскими взаимозаменяемыми могут быть как меньший посадочный!!! размер 235/65 R17 и 255/60 R17 так и эквивалентный, но более узкий 235/60 R18, так и 255/50 R19, 285/45 R19 в 19м посадочном диаметре, 245/45 R20, 275/40 R20 и даже 315/35 R20, и даже 22 дюймовые аналоги 285/30 R22. Так и для большинства легковых авто среднего класса суперпопулярный размер 205/55 R16 заменяем размерами 195/65 R15 и 205/60 R15 при уменьшении посадочного диаметра шины, 195/60 R16 и 225/50 R16 при необходимости изменения ширины покрышки, 205/50 R17 и 225/45 R17 для понижения профиля шин, 225/40 R18 и 255/35 R18 для еще более спортивного поведения автомобиля и 225/35 R19 и соответственно 255/30 R19 для самых экстремалов. Более широкие варианты покрышек предназначены для подбора шины для заднеприводных автомобилей Мерседес и БМВ.
Внутренний, то есть посадочный диаметр и высота профиля шины являются обратно пропорциональными друг другу и самыми “гибкими” при подборе величинами. Чем ниже профиль, тем больше посадочный диаметр диска и наоборот. Главное – сохранение внешнего или полного диаметра колеса, что и позволяет вычислить шинный калькулятор. При уменьшении диаметра шины необходимо учитывать очень важный момент – диски меньшего диаметра могут просто не налезть на тормозной суппорт. Обязательно необходима примерка именно на ваш автомобиль (очень актуально для немцев даже при увеличении диаметра), так как очень часто бывали случаи, когда диск не только не одевается из-за большого тормозного механизма вообще, но и не может прислониться к ступице из-за недостаточно выпуклой формы диска. Даже если у товарища на такой же чуть более старой или менее мощной машине все подошло, нельзя быть уверенным, что подойдет на вашей. Автопроизводители не дремлют:) и постоянно суют палки в колеса, внося изменения в конструкцию авто:)
Чем ниже профиль, тем больше посадочный диаметр диска и наоборот. Главное – сохранение внешнего или полного диаметра колеса, что и позволяет вычислить шинный калькулятор. При уменьшении диаметра шины необходимо учитывать очень важный момент – диски меньшего диаметра могут просто не налезть на тормозной суппорт. Обязательно необходима примерка именно на ваш автомобиль (очень актуально для немцев даже при увеличении диаметра), так как очень часто бывали случаи, когда диск не только не одевается из-за большого тормозного механизма вообще, но и не может прислониться к ступице из-за недостаточно выпуклой формы диска. Даже если у товарища на такой же чуть более старой или менее мощной машине все подошло, нельзя быть уверенным, что подойдет на вашей. Автопроизводители не дремлют:) и постоянно суют палки в колеса, внося изменения в конструкцию авто:)
Увеличение ширины покрышки ведет к лучшей управляемости в поворотах и возможности прохождения этих поворотов на большей скорости, уменьшению тормозного пути на любом покрытии (узкие шины для зимы – полное заблуждение особенно на твердом покрытии). А недостатками широких шин являются повышенный расход топлива, резкие реакции на колеи и прочие неровности на дорогах. Увеличение диаметра колесного диска ведет к ухудшению комфорта и повышенной жесткости, но позволяет сделать машину более спортивной за счет меньшего увода (смещения) протектора от ободов диска в поперечном направлении в резких и скоростных поворотах.
А недостатками широких шин являются повышенный расход топлива, резкие реакции на колеи и прочие неровности на дорогах. Увеличение диаметра колесного диска ведет к ухудшению комфорта и повышенной жесткости, но позволяет сделать машину более спортивной за счет меньшего увода (смещения) протектора от ободов диска в поперечном направлении в резких и скоростных поворотах.
Увеличение ширины покрышки всегда осуществляется автовладельцем на свой страх и риск и не может служить поводом или причиной для предъявления претензий сотрудникам магазина или техцентра. Вся ответственность в данном поступке лежит на автовладельце или его представителе.
Следует учесть также, что индекс скорости и индекс нагрузки шины обычно выше у покрышек большего посадочного диаметра и более низкого профиля, поэтому увеличение посадочного диаметра диска только положительно сказывается на управляемости и качестве сцепления с дорожным покрытием, ущемляя только комфорт от тряски в автомобиле.
Данный калькулятор подбора дисков служит, чтобы клиент сам мог осуществить подбор дисков под шины или шин под диски по ширине.
Ширина колесного диска может варьироваться для одних и тех же покрышек в отличии от диаметра диска. Это вызвано тем, что конструкция корда не мешает резиновым боковинам покрышки располагаться чуть шире или уже в допустимых пределах (например на BMW E39 производитель ставил 225 шины на 16 диски шириной 7 дюймов, а на Mercedes W210 в тоже самое время завод устанавливал 215 шины на 16 диски шириной 7,5 дюймов при одинаковой высоте профиля 55%; при этом параметры 17 шин и дисков на этих двух моделях совпадали полностью). А вот стальные бортовые кольца, определяющие диаметр и отвечающие за плотную и герметичную посадку шины на диске, таких вольностей не допускают.
При монтаже на слишком широкие для данных шин диски покрышка встанет на них “домиком” – это очень не красиво (в разрезе она будет смотреться как трапеция), диск будет шире резины и будет получать повреждения о каждый бордюр или камень на дороге. А главное, уменьшится площадь соприкосновения плоскости протектора с дорогой.
При установке же шин на слишком узкие для них диски покрышки изогнутся по всей своей длине как капюшон у кобры. Это совсем недопустимо. Помимо уменьшенного пятна контакта и повышенного износа центральной части протектора шины неизбежно начнется необратимая деформация стального корда покрышки.
При подборе колес на авто нужно обязательно учитывать вылет диска, то есть смещение относительно ступицы наружу автомобиля, что не позволяет сделать данный дисковый калькулятор. При установке новых дисков необходима обязательная их примерка до монтажа шин. Это недорогая, но очень ответственная операция по принципу семь раз отмерь даже более необходима, чем примерка обуви вашего размера перед покупкой в магазине, так как металл не разносится со временем, а должен подходить сразу. Многообразие формы колесных суппортов современных автомобилей и внутренней поверхности всего разнообразного ассортимента колесных дисков не позволяет полагаться ни на один каталог.
При изменении полного диаметра колеса изменятся показания спидометра в том процентном соотношении, что показывает наш автомобильный шинный калькулятор. При увеличении диаметра более чем на 1,5% скорее всего станет невозможным использование автомобильных цепей для снежного или грязевого покрытия.
При увеличении диаметра более чем на 1,5% скорее всего станет невозможным использование автомобильных цепей для снежного или грязевого покрытия.
Подбор шин большей ширины данным калькулятором не отменяет необходимости обязательной примерки дисков, так как часты случаи использования дисков с неправильным вылетом, из-за чего все колесо может выступать наружу и цеплять за крыло на выбоинах или ухабах, или просто забрызгивать авто грязью. Рекомендуется также проверка полностью загруженного авто до выезда из техцентра при установке широких шин или уменьшении значения вылета дисков для исключения трения о детали подвески или крылья машины. Шипованные шины требуют двойного внимания при эксперименте с подбором, так как способны прямо как фреза очень быстро распилить любое крыло.
Шиномонтаж в «Автобам Запад» (репортаж)
Устройство вакуумной фильтрации — PatentDB.
 ru
ruУстройство вакуумной фильтрации
Иллюстрации
Показать всеИзобретение предназначено для фильтрации. Устройство для фильтрации пульпы с помощью вакуума содержит корпус, фильтровальную ткань, на которую подается пульпа и которая перемещается, по существу, горизонтально относительно корпуса устройства, вакуумный лоток, расположенный под фильтровальной тканью и выполняющий возвратно-поступательные движения в направлении перемещения фильтровальной ткани, вакуумные шланги, обладающие эластичностью, причем концы, по меньшей мере, двух вакуумных шлангов присоединены к вакуумному лотку таким образом, чтобы проходить в противоположных направлениях относительно середины вакуумного лотка, при этом другие их концы присоединены к вакуумному устройству, которое прикладывает вакуум к пульпе через фильтровальную ткань и вакуумный лоток. Технический результат: обеспечение надежности устройства.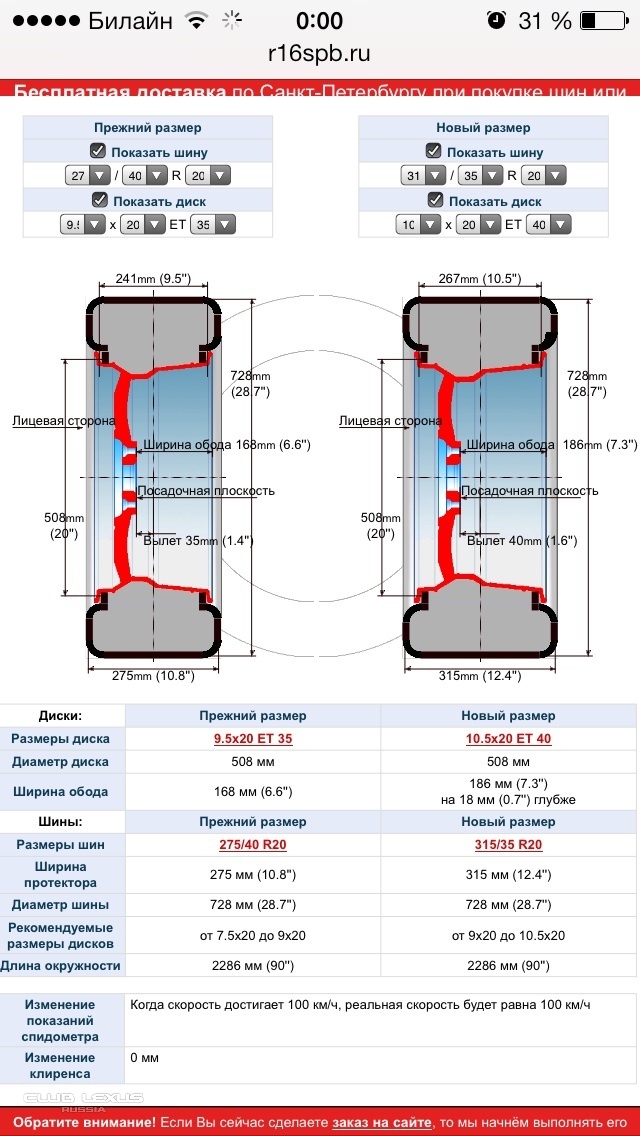 4 з.п. ф-лы, 5 ил.
4 з.п. ф-лы, 5 ил.
Реферат
Предпосылки к созданию изобретения
Область техники, к которой относится изобретение
Настоящее изобретение относится к устройству вакуумной фильтрации, в котором пульпа, поступающая на фильтровальную ткань, фильтруется с использованием вакуумных лотков для применения вакуума, где фильтровальная ткань может перемещаться, по существу, по горизонтали, а вакуумные лотки помещаются под этой фильтровальной тканью.
Приоритет заявлен по японской патентной заявке №2004-089346, поданной 25 марта 2004 г., содержание которой включено сюда в качестве ссылки.
Описание предшествующего уровня техники
Пример устройства вакуумной фильтрации такого типа представлен в ссылке 1 (Нерассмотренная японская патентная заявка, первая публикация, №S62-79817) и в ссылке 2 (Нерассмотренная японская патентная заявка, первая публикация, №Н07-148404). В этих ссылках предложены устройства, в которых бесконечная имеющая форму ленты фильтровальная ткань подвешена и расположена вокруг множества роликов, размещенных на корпусе устройства, причем эта фильтровальная ткань может непрерывно перемещаться по кругу. В этих устройствах в той части перемещающейся фильтровальной ткани, которая перемещается по горизонтали над верхней частью корпуса устройства, множество вакуумных лотков, помещенных под этой фильтровальной тканью, может выполнять возвратно-поступательные движения в направлении перемещения этой фильтровальной ткани с заданным шагом. Эти вакуумные лотки соединены каждый с вакуумным устройством посредством соединительных фитингов. Когда вакуумные лотки продвигаются вперед, в направлении перемещения фильтровальной ткани, пульпа, поступающая на фильтровальную ткань, фильтруется благодаря вакууму, который обеспечивается через фильтровальную ткань посредством вакуумного устройства. В отличие от этого, когда вакуумные лотки достигают конца шага, вакуум сбрасывается, и вакуумные лотки возвращаются в исходное положение, после чего операция, включая всасывание и фильтрацию, повторяется во время повторного перемещения вакуумных лотков вперед.
В этих ссылках предложены устройства, в которых бесконечная имеющая форму ленты фильтровальная ткань подвешена и расположена вокруг множества роликов, размещенных на корпусе устройства, причем эта фильтровальная ткань может непрерывно перемещаться по кругу. В этих устройствах в той части перемещающейся фильтровальной ткани, которая перемещается по горизонтали над верхней частью корпуса устройства, множество вакуумных лотков, помещенных под этой фильтровальной тканью, может выполнять возвратно-поступательные движения в направлении перемещения этой фильтровальной ткани с заданным шагом. Эти вакуумные лотки соединены каждый с вакуумным устройством посредством соединительных фитингов. Когда вакуумные лотки продвигаются вперед, в направлении перемещения фильтровальной ткани, пульпа, поступающая на фильтровальную ткань, фильтруется благодаря вакууму, который обеспечивается через фильтровальную ткань посредством вакуумного устройства. В отличие от этого, когда вакуумные лотки достигают конца шага, вакуум сбрасывается, и вакуумные лотки возвращаются в исходное положение, после чего операция, включая всасывание и фильтрацию, повторяется во время повторного перемещения вакуумных лотков вперед.
Соединительные фитинги для устройства вакуумной фильтрации такого типа обычно снабжены вакуумными трубами, которые подсоединены к нижней поверхности вакуумных лотков, и вакуумными шлангами, имеющими первый конец, присоединенный к вакуумной трубе, и второй конец, присоединенный к вакуумному устройству.
Вакуумная труба непосредственно присоединена к нижней поверхности вакуумного лотка. Вакуумный шланг изготовлен из эластичного (или гибкого) материала. Второй конец вакуумного шланга расположен на одном конце фильтровальной ткани в поперечном направлении и присоединен к боковой поверхности вакуумного устройства. В отличие от этого, первый конец вакуумного шланга расположен в средней части вакуумного лотка в поперечном направлении фильтровальной ткани. Вакуумные шланги расположены ниже вакуумных лотков, и изогнуты для того, чтобы обеспечить возвратно-поступательное движение вакуумных лотков. В случае крупномасштабного устройства вакуумной фильтрации, в котором используются вакуумные лотки с большой длиной в поперечном направлении, применение множества вакуумных трубок, расположенных через определенные интервалы в поперечном направлении, обеспечивает равномерное действие вакуума из вакуумного лотка, приложенного к пульпе, в поперечном направлении с надежным отбором фильтрата из пульпы.
В случае крупномасштабного устройства вакуумной фильтрации, в котором используются вакуумные лотки с большой длиной в поперечном направлении, применение множества вакуумных трубок, расположенных через определенные интервалы в поперечном направлении, обеспечивает равномерное действие вакуума из вакуумного лотка, приложенного к пульпе, в поперечном направлении с надежным отбором фильтрата из пульпы.
Однако в устройстве вакуумной фильтрации такого типа, из-за того что эластичные вакуумные шланги, описанные выше, расположены в изогнутом состоянии, сопровождая возвратно-поступательное движение вакуумных лотков, восстанавливающие силы, которые служат для восстановления деформированных вакуумных шлангов в их первоначальное состояние, воздействуют на вакуумные лотки. Поэтому, в связи с тем что вакуумные лотки подвергаются воздействию усилий, ориентированных поперек направления их движения, оказывается трудно поддерживать прямолинейную стабильность вакуумных лотков.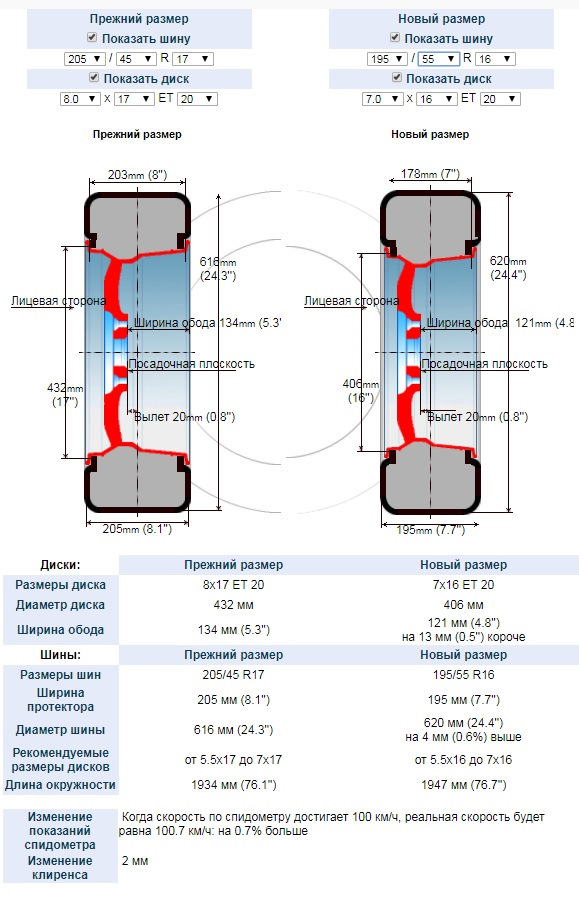
Когда восстанавливающие силы воздействуют таким образом на вакуумные лотки в случае, например, когда вакуумные лотки опираются на конструкцию, в которой предусмотрены опорные колеса, находящиеся в контакте с обеими сторонами вакуумных лотков, начиная от дна в поперечном направлении, возможен неравномерный износ, который вызывает наклон вакуумных лотков. Неравномерный износ показывает состояние, при котором величина износа опорных валков и т.п. различается на каждой стороне в поперечном направлении. Когда вакуумные лотки наклонены, существует опасение того, что перемещение вакуумных лотков будет затруднено из-за неравномерного движения фильтровальной ткани. Кроме того, в случае, когда, например, на боковой поверхности вакуумных лотков предусмотрена направляющая, которая ограничивает вакуумные лотки в направлении перемещения в связи с описанным выше наклоном, возникает ситуация, при которой боковая поверхность вакуумных лотков скребется о направляющую, и таким образом возникает опасение, что препятствие перемещению вакуумных лотков возникает еще легче.
В частности, в случае крупномасштабного устройства вакуумной фильтрации вакуумные шланги должны иметь большой диаметр, поскольку необходимо создавать вакуум, которые равномерно будут прилагаться к пульпе в поперечном направлении вакуумного лотка, и поскольку фильтрат, отбираемый из пульпы, может быть получен в большом количестве. Однако, поскольку вакуумные шланги изогнуты, описанная выше проблема, при которой перемещение вакуумных лотков может быть затруднено, становится серьезнее, поскольку усилие, воздействующее на вакуумные лотки, возрастает из-за того, что увеличивается диаметр вакуумных шлангов. Кроме того, когда диаметр вакуумных шлангов возрастает таким образом, то радиус кривизны вакуумных шлангов становится больше. Поэтому существует проблема, заключающаяся в том, что обеспечение пространства, вмещающего такие вакуумные шланги, становится затруднительным при размещении вакуумных шлангов в изогнутом состоянии, описанном выше.
Кроме того, в конструкции, в которой множество вакуумных лотков размещено в направлении перемещения, и соединительные фитинги размещены на каждом из этих вакуумных лотков, интервал между соседними вакуумными шлангами становится небольшим в направлении перемещения в связи с увеличением диаметра вакуумных шлангов. Таким образом, существует опасение, что возрастет длительность технического обслуживания вакуумных лотков. В отличие от этого, если увеличивается количество соединительных фитингов и увеличивается количество вакуумных шлангов для того, чтобы избежать увеличения диаметра вакуумных шлангов, возникает проблема, связанная с переплетением этих шлангов между собой и переплетением вакуумных шлангов и опорных колес.
Таким образом, существует опасение, что возрастет длительность технического обслуживания вакуумных лотков. В отличие от этого, если увеличивается количество соединительных фитингов и увеличивается количество вакуумных шлангов для того, чтобы избежать увеличения диаметра вакуумных шлангов, возникает проблема, связанная с переплетением этих шлангов между собой и переплетением вакуумных шлангов и опорных колес.
С учетом описанных выше проблем задачей настоящего изобретения является создание устройства вакуумной фильтрации, при котором перемещение вакуумных лотков является благоприятным и надежным.
Сущность изобретения
Поставленная задача достигается посредством создания устройства вакуумной фильтрации, описанного ниже. На корпусе устройства помещена фильтровальная ткань, которая может перемещаться, по существу, по горизонтали. Под этой фильтровальной тканью размещены вакуумные лотки, которые соединены с вакуумным устройством посредством соединительных фитингов, причем эти вакуумные лотки могут выполнять возвратно-поступательные движения в направлении перемещения фильтровальной ткани. Предусмотрено множество соединительных фитингов, каждый из этих соединительных фитингов образует эластичный вакуумный шланг. Эти вакуумные шланги расположены таким образом, что могут проходить в противоположном относительно друг друга направлении относительно средней части вакуумных лотков. В этом устройстве обеспечивается возможность фильтрации пульпы, поступающей на фильтровальную ткань, путем приложения вакуума сквозь фильтровальную ткань вакуумных лотков.
Предусмотрено множество соединительных фитингов, каждый из этих соединительных фитингов образует эластичный вакуумный шланг. Эти вакуумные шланги расположены таким образом, что могут проходить в противоположном относительно друг друга направлении относительно средней части вакуумных лотков. В этом устройстве обеспечивается возможность фильтрации пульпы, поступающей на фильтровальную ткань, путем приложения вакуума сквозь фильтровальную ткань вакуумных лотков.
Согласно настоящему изобретению применяется множество соединительных фитингов, и вакуумные шланги каждого из этих соединительных фланцев расположены таким образом, что могут проходить в противоположном относительно друг друга направлении относительно средней части вакуумных лотков в поперечном направлении. Таким образом, даже в случае, когда на вакуумные лотки воздействуют восстанавливающие силы, когда восстанавливающие силы должны восстанавливать каждый из вакуумных шлангов, который деформируется при возвратно-поступательном движении вакуумных лотков, в его первоначальное состояние, обеспечивается возможность взаимной нейтрализации этих сил на вакуумных лотках. В связи с этим существует возможность ограничивать до минимума силы, действующие на вакуумные лотки путем восстановления усилий, действующих на вакуумные лотки в направлении, поперечном относительно направления перемещения, и существует возможность ограничить вероятность возникновения ситуации, при которой перемещение этих вакуумных лотков будет затруднено, и таким образом можно обеспечить прямолинейную стабильность вакуумных лотков. Например, даже в случае, в котором для поддержки вакуумных лотков предусмотрены опорные колеса и т.п., которые соприкасаются снизу с обоими концами вакуумных лотков в поперечном направлении, возникает возможность ограничить неравномерный износ, при котором величина износа опорных колес и т.п. различается на каждом конце в поперечном направлении, существует возможность ограничить наклон вакуумных лотков, и таким образом можно ограничить вероятность возникновения ситуации, при которой фильтровальная ткань легко перемещается неравномерно и перемещение вакуумного лотка будет затруднено.
В связи с этим существует возможность ограничивать до минимума силы, действующие на вакуумные лотки путем восстановления усилий, действующих на вакуумные лотки в направлении, поперечном относительно направления перемещения, и существует возможность ограничить вероятность возникновения ситуации, при которой перемещение этих вакуумных лотков будет затруднено, и таким образом можно обеспечить прямолинейную стабильность вакуумных лотков. Например, даже в случае, в котором для поддержки вакуумных лотков предусмотрены опорные колеса и т.п., которые соприкасаются снизу с обоими концами вакуумных лотков в поперечном направлении, возникает возможность ограничить неравномерный износ, при котором величина износа опорных колес и т.п. различается на каждом конце в поперечном направлении, существует возможность ограничить наклон вакуумных лотков, и таким образом можно ограничить вероятность возникновения ситуации, при которой фильтровальная ткань легко перемещается неравномерно и перемещение вакуумного лотка будет затруднено. Кроме того, например, даже в случае размещения на боковой поверхности вакуумных лотков направляющих, которые ограничивают вакуумные лотки в направлении перемещения, поскольку становится возможным ограничение наклона вакуумных лотков, описанного выше, имеется возможность ограничить ситуацию, в которой направляющие скребутся о боковые поверхности, и имеется возможность ограничить препятствия перемещению вакуумных лотков. Кроме того, поскольку применяется множество соединительных фитингов, даже в случае, если диаметр вакуумных шлангов этих соединительных фитингов небольшой, имеется возможность поддерживать воздействие вакуума, которое прилагается к пульпе из вакуумных лотков. Если диаметр вакуумных шлангов сделать небольшим, восстанавливающее усилие вакуумных шлангов становится также небольшим, благодаря этому становится возможным дополнительно и надежно ограничить возможность возникновения ситуации, при которой возникают препятствия перемещению вакуумных лотков. Кроме того, если диаметр вакуумных шлангов сделан небольшим, возникает возможность экономии пространства под вакуумными лотками, и можно обеспечить сокращение затрат времени на техническое обслуживание этой части.
Кроме того, например, даже в случае размещения на боковой поверхности вакуумных лотков направляющих, которые ограничивают вакуумные лотки в направлении перемещения, поскольку становится возможным ограничение наклона вакуумных лотков, описанного выше, имеется возможность ограничить ситуацию, в которой направляющие скребутся о боковые поверхности, и имеется возможность ограничить препятствия перемещению вакуумных лотков. Кроме того, поскольку применяется множество соединительных фитингов, даже в случае, если диаметр вакуумных шлангов этих соединительных фитингов небольшой, имеется возможность поддерживать воздействие вакуума, которое прилагается к пульпе из вакуумных лотков. Если диаметр вакуумных шлангов сделать небольшим, восстанавливающее усилие вакуумных шлангов становится также небольшим, благодаря этому становится возможным дополнительно и надежно ограничить возможность возникновения ситуации, при которой возникают препятствия перемещению вакуумных лотков. Кроме того, если диаметр вакуумных шлангов сделан небольшим, возникает возможность экономии пространства под вакуумными лотками, и можно обеспечить сокращение затрат времени на техническое обслуживание этой части. В данном случае вакуумные шланги могут быть расположены симметрично относительно средней части вакуумных лотков в поперечном направлении. В этом случае восстанавливающие усилия, связанные с каждым из вакуумных шлангов и воздействующие на вакуумные лотки, могут быть надежно устранены, и возникает возможность дополнительно улучшить прямолинейную стабильность вакуумных лотков.
В данном случае вакуумные шланги могут быть расположены симметрично относительно средней части вакуумных лотков в поперечном направлении. В этом случае восстанавливающие усилия, связанные с каждым из вакуумных шлангов и воздействующие на вакуумные лотки, могут быть надежно устранены, и возникает возможность дополнительно улучшить прямолинейную стабильность вакуумных лотков.
Можно также присоединять вакуумные шланги к вакуумным лоткам снизу в положении, при котором вакуумные шланги изогнуты таким образом, чтобы образовать выпуклость, обращенную вверх или вниз по направлению перемещения фильтровальной ткани. В этом случае существует возможность протянуть вакуумные шланги в направлении вдоль поверхности фильтровальной ткани или вакуумных лотков и появляется возможность гарантировать образование пространства под вакуумными лотками, благодаря чему становится возможным ограничить затраты времени на техническое обслуживание этой части.
Вакуумные шланги могут быть присоединены к вакуумным лоткам снизу таким образом, чтобы они оставались отделены друг от друга, причем средняя часть вакуумных лотков в поперечном направлении расположена между вакуумными шлангами. В этом случае под вакуумными лотками благодаря тому, что вакуумные шланги не пересекаются между собой, появляется возможность надежно предотвратить пересечение между каждым из вакуумных шлангов и в то же время можно гарантировать наличие даже большего пространства под вакуумными лотками.
В этом случае под вакуумными лотками благодаря тому, что вакуумные шланги не пересекаются между собой, появляется возможность надежно предотвратить пересечение между каждым из вакуумных шлангов и в то же время можно гарантировать наличие даже большего пространства под вакуумными лотками.
Можно также конфигурировать множество вакуумных лотков, разместив их в направлении перемещения фильтровальной ткани или присоединив вакуумные шланги каждого из этих вакуумных лотков снизу. В этом случае, поскольку вакуумные шланги имеют небольшой диаметр из-за множественного присоединения соединительных фитингов, пересечение между соседними вакуумными шлангами в направлении перемещения ограничивается. Поэтому можно обеспечить уменьшение количества и типов деталей, которые образуют это устройство вакуумной фильтрации, и можно обеспечить как снижение стоимости этого устройства, так и снижение затрат на эксплуатацию и обслуживание этого устройства.
Краткое описание чертежей
На фиг.1 изображен вид сбоку всей конструкции варианта реализации устройства вакуумной фильтрации согласно настоящему изобретению;
на фиг.2 изображен вид сверху устройства вакуумной фильтрации, показанного на фиг.1;
на фиг.3 изображен поперечный разрез вдоль линии А-А устройства вакуумной фильтрации, показанного на фиг.1;
на фиг.4 изображен участок В в увеличенном масштабе устройства вакуумной фильтрации, показанного на фиг.3;
на фиг.5 изображен поперечный разрез вдоль линии С-С устройства вакуумной фильтрации, показанного на фиг.3.
Подробное описание изобретения
На фиг.1-5 показан вариант реализации настоящего изобретения. В устройстве вакуумной фильтрации согласно настоящему изобретению на корпусе 10 устройства подвешена и последовательно обведена вокруг роликов 12 бесконечная имеющая форму ленты фильтровальная ткань 11. Верхний участок этой фильтровальной ткани 11 расположен таким образом, чтобы проходить в горизонтальном направлении, и этот участок служит фильтрующей частью 11А. Кроме того, среди множества роликов 12 ролик 12А, на котором лежит и который обмотан фильтровальной тканью 11 с одного конца этой фильтрующей части 11А (с правой стороны на фиг.1), соединен с приводным устройством (не показано). Ролик 12А вращается приводным устройством по часовой стрелке, и таким образом фильтровальная ткань 11 также непрерывно вращается поверх роликов 12. Сопровождая это вращение, фильтрующая часть 11А фильтровальной ткани 11 непрерывно перемещается в направлении перемещения, показанном стрелкой F.
В устройстве вакуумной фильтрации согласно настоящему изобретению на корпусе 10 устройства подвешена и последовательно обведена вокруг роликов 12 бесконечная имеющая форму ленты фильтровальная ткань 11. Верхний участок этой фильтровальной ткани 11 расположен таким образом, чтобы проходить в горизонтальном направлении, и этот участок служит фильтрующей частью 11А. Кроме того, среди множества роликов 12 ролик 12А, на котором лежит и который обмотан фильтровальной тканью 11 с одного конца этой фильтрующей части 11А (с правой стороны на фиг.1), соединен с приводным устройством (не показано). Ролик 12А вращается приводным устройством по часовой стрелке, и таким образом фильтровальная ткань 11 также непрерывно вращается поверх роликов 12. Сопровождая это вращение, фильтрующая часть 11А фильтровальной ткани 11 непрерывно перемещается в направлении перемещения, показанном стрелкой F.
Устройство 13 подачи пульпы, которое подает пульпу S на фильтровальную ткань 11, размещено на другом конце фильтрующей части 11А фильтровальной ткани 11 (с левой стороны на фиг. 1), т.е. сверху и перед направлением перемещения F. Пульпа S, которая поступает из этого устройства 13 подачи пульпы на фильтровальную ткань 11 над вакуумным лотком 16, фильтруется под воздействием вакуума, прилагаемого вакуумным лотком 16 через фильтровальную ткань 11 во время перенесения в направлении перемещения F благодаря перемещению фильтрующей части 11А. Кроме того, над фильтрующей частью 11А со стороны, противоположной устройству 13 подачи пульпы в направлении перемещения F, предусмотрено устройство 21 промывки «пирога», которое подает на фильтровальную ткань 11 промывочную жидкость L. Промывочная жидкость L поступает из этого устройства 21 промывки «пирога» на пульпу S, которая подвергается вакуумной фильтрации. На ролике 12А, который осуществляет привод фильтровальной ткани 11, предусмотрен нож 14, который отделяет и удаляет от фильтровальной ткани обезвоженный пирог пульпы S, фильтрация и обезвоживание которого закончены. Кроме того, устройство 15 промывки фильтровальной ткани, которое промывает фильтровальную ткань 11 после отделения пульпы S ножом 14, размещено под одним концом фильтрующей части 11А фильтровальной ткани 11 (с правой стороны на фиг.
1), т.е. сверху и перед направлением перемещения F. Пульпа S, которая поступает из этого устройства 13 подачи пульпы на фильтровальную ткань 11 над вакуумным лотком 16, фильтруется под воздействием вакуума, прилагаемого вакуумным лотком 16 через фильтровальную ткань 11 во время перенесения в направлении перемещения F благодаря перемещению фильтрующей части 11А. Кроме того, над фильтрующей частью 11А со стороны, противоположной устройству 13 подачи пульпы в направлении перемещения F, предусмотрено устройство 21 промывки «пирога», которое подает на фильтровальную ткань 11 промывочную жидкость L. Промывочная жидкость L поступает из этого устройства 21 промывки «пирога» на пульпу S, которая подвергается вакуумной фильтрации. На ролике 12А, который осуществляет привод фильтровальной ткани 11, предусмотрен нож 14, который отделяет и удаляет от фильтровальной ткани обезвоженный пирог пульпы S, фильтрация и обезвоживание которого закончены. Кроме того, устройство 15 промывки фильтровальной ткани, которое промывает фильтровальную ткань 11 после отделения пульпы S ножом 14, размещено под одним концом фильтрующей части 11А фильтровальной ткани 11 (с правой стороны на фиг.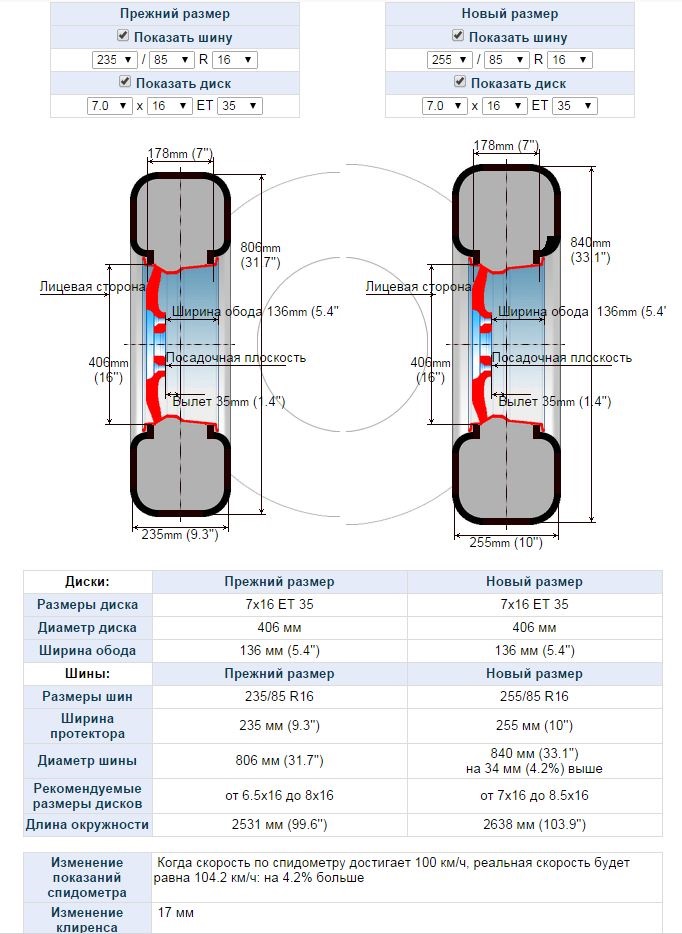 1), т.е. по направлению вперед относительно перемещения F. Здесь вакуумный лоток 16 соединен с вакуумным устройством (не показано) соединительным фитингом 20. Вакуум прилагается посредством вакуумного устройства через фильтровальную ткань 11 к пульпе S, которую подают на фильтровальную ткань 11. Множество вакуумных лотков 16 размещено под фильтрующей частью 11А фильтровальной ткани 11, начиная от позиции непосредственно под устройством 13 подачи пульпы и по направлению вперед относительно перемещения F. Соединительные фитинги 20 присоединены к каждому из этих вакуумных лотков 16 снизу.
1), т.е. по направлению вперед относительно перемещения F. Здесь вакуумный лоток 16 соединен с вакуумным устройством (не показано) соединительным фитингом 20. Вакуум прилагается посредством вакуумного устройства через фильтровальную ткань 11 к пульпе S, которую подают на фильтровальную ткань 11. Множество вакуумных лотков 16 размещено под фильтрующей частью 11А фильтровальной ткани 11, начиная от позиции непосредственно под устройством 13 подачи пульпы и по направлению вперед относительно перемещения F. Соединительные фитинги 20 присоединены к каждому из этих вакуумных лотков 16 снизу.
Как показано на фиг.3, каждый из этих вакуумных лотков 16 имеет плоскую часть 16А, которая проходит вдоль поверхности фильтровальной ткани 11 и находится в тесном контакте с этой фильтровальной тканью 11, стенки 16В, которые проходят вверх от двух кромок плоской части 16А в поперечном направлении наружу, так что обе расходятся, и армирующую трубу 17, прикрепленную к плоской части 16А снизу и проходящую в поперечном направлении.
Далее, среди направлений вдоль поверхности фильтровальной ткани 11, направление, поперечное относительно направления перемещения F, обозначено выражением «поперечное направление вакуумных лотков 16».
Как показано на фиг.4, плоская часть 16А является слоистым телом, в котором пластина 16С и сетчатое тело 16D размещены слоями в этой последовательности снизу. Сетчатое тело 16D имеет множество сквозных отверстий 16F, выполненных на поверхности. Фильтровальная ткань 11 находится в тесном контакте с поверхностной стороной сетчатого тела 16D. Как показано на фиг.2, множество первых соединительных отверстий 16Е просверлено в поверхности пластины 16С таким образом, чтобы они были отделены промежутками в направлении, поперечном относительно фильтровальной ткани 11 (в направлении по вертикали на фиг.2). Среди этих первых соединительных отверстий 16Е половина отверстий просверлена таким образом, чтобы находиться на одинаковых расстояниях друг от друга в поперечном направлении на одной стороне середины фильтровальной ткани 11 в поперечном направлении, а остальная половина отверстий просверлена таким образом, чтобы находиться на одинаковых расстояниях друг от друга на другой стороне средней части в поперечном направлении.
Армирующие трубы 17 прикреплены к каждой из пластин 16С снизу и выполнены так, чтобы проходить в поперечном направлении относительно фильтровальной ткани 11, то есть в поперечном направлении относительно вакуумного лотка 16. Как показано на фиг.5, армирующая труба 17 имеет прямоугольное поперечное сечение, а внутри нее имеется внутренняя полость 17С, которая проходит в поперечном направлении. Кроме того, вторые соединительные отверстия 17А, диаметр которых меньше диаметра первых соединительных отверстий 16Е, просверлены в местах, совмещенных с местами, где в пластине 16С просверлены первые соединительные отверстия 16С. Верхняя сторона пластины 16С и внутренняя полость 17С армирующей трубы 17 сообщаются благодаря вторым соединительным отверстиям 17А и первым соединительным отверстиям 16Е. Как показано на фиг.2, в местах между самой внутренней парой вторых соединительных отверстий 17А в поперечном направлении на нижней поверхности армирующей трубы 17 в противоположных местах просверлены два третьих соединительных отверстия 17В, а середина фильтровальной ткани 11 в поперечном направлении расположена между ними. Как показано на фиг.2 и фиг.5, между группой соседних армирующих труб 17 в направлении перемещения F фильтровальной ткани предусмотрены армирующие элементы (например, элементы типа швеллеров) 17D, которые проходят в направлении перемещения F фильтровальной ткани 11. Эти элементы типа швеллеров 17D скреплены с нижней поверхностью пластин 16С и боковой поверхностью армирующих труб 17.
Как показано на фиг.2 и фиг.5, между группой соседних армирующих труб 17 в направлении перемещения F фильтровальной ткани предусмотрены армирующие элементы (например, элементы типа швеллеров) 17D, которые проходят в направлении перемещения F фильтровальной ткани 11. Эти элементы типа швеллеров 17D скреплены с нижней поверхностью пластин 16С и боковой поверхностью армирующих труб 17.
Как показано на фиг.3 и 5, соединительный фитинг 20 снабжен вакуумной трубой 20A, которая соединена с третьим соединительным отверстием 17В в армирующей трубе 17 и проходит вниз, а также вакуумным шлангом 20В, один конец которого соединен с нижним концом вакуумной трубы 20A, а другой конец соединен с вакуумным устройством (не показано). Вакуумная труба 20A образована жесткой трубой, выполненной, например, из нержавеющей стали. Вакуумный шланг 20В выполнен из эластичного или гибкого материала, такого как этиленпропиленовый каучук.
Эти соединительные фитинги 20 размещены по несколько штук (в данном варианте реализации два) на одном вакуумном лотке 16. Под вакуумным лотком 16 эти соединительные фитинги 20 расположены напротив друг друга таким образом, чтобы находиться симметрично в поперечном направлении относительно середины фильтровальной ткани 11, когда средняя часть фильтровальной ткани 11 расположена в поперечном направлении между ними. Как показано на фиг.2, каждый из вакуумных шлангов 20В проходит в поперечном направлении относительно фильтровальной ткани 11 от средней части фильтровальной ткани 11 таким образом, чтобы отделяться друг от друга. Кроме того, как показано на фиг.3, участок вакуумных шлангов 20В между одним концом и другим концом установлен таким образом, чтобы иметь возможность скользить по имеющим форму рельсов опорным элементам 20С, которые проходят в направлении перемещения F. Этот участок, по существу, параллелен поверхности фильтровальной ткани 11 под вакуумными лотками 16 и искривлен таким образом, чтобы выпуклость была обращена вперед по направлению F фильтровальной ткани 11. Путем установки таким образом вакуумных шлангов 20В они удерживаются в положении, обеспечивающем возвратно-поступательное движение в направлении перемещения F.
Под вакуумным лотком 16 эти соединительные фитинги 20 расположены напротив друг друга таким образом, чтобы находиться симметрично в поперечном направлении относительно середины фильтровальной ткани 11, когда средняя часть фильтровальной ткани 11 расположена в поперечном направлении между ними. Как показано на фиг.2, каждый из вакуумных шлангов 20В проходит в поперечном направлении относительно фильтровальной ткани 11 от средней части фильтровальной ткани 11 таким образом, чтобы отделяться друг от друга. Кроме того, как показано на фиг.3, участок вакуумных шлангов 20В между одним концом и другим концом установлен таким образом, чтобы иметь возможность скользить по имеющим форму рельсов опорным элементам 20С, которые проходят в направлении перемещения F. Этот участок, по существу, параллелен поверхности фильтровальной ткани 11 под вакуумными лотками 16 и искривлен таким образом, чтобы выпуклость была обращена вперед по направлению F фильтровальной ткани 11. Путем установки таким образом вакуумных шлангов 20В они удерживаются в положении, обеспечивающем возвратно-поступательное движение в направлении перемещения F. Скользящие элементы 20D прикреплены на участках вакуумных шлангов 20В, которые соприкасаются с опорными элементами 20С. Эти скользящие элементы 20С выполнены из материала, обладающего низким коэффициентом трения с опорными элементами 20С и имеющего повышенную износостойкость, такого как полиэтилен со сверхвысокой молекулярной массой.
Скользящие элементы 20D прикреплены на участках вакуумных шлангов 20В, которые соприкасаются с опорными элементами 20С. Эти скользящие элементы 20С выполнены из материала, обладающего низким коэффициентом трения с опорными элементами 20С и имеющего повышенную износостойкость, такого как полиэтилен со сверхвысокой молекулярной массой.
В данном случае два соединительных фитинга 20 расположены напротив друг друга в поперечном направлении и размещены таким образом, что каждая из их вакуумных труб 20А расположена, по существу, идентично относительно направления перемещения F. Кроме того, с обеих сторон корпуса 10 устройства в поперечном направлении размещены всасывающие трубы большого диаметра (не показаны), которые проходят параллельно направлению перемещения F. Другие концы вакуумных шлангов 20В двух соединительных фитингов 20 соединены каждый со всасывающими трубами. Посредством этих всасывающих труб соединительные фитинги соединены с вакуумным устройством.
В описанном выше устройстве вакуумной фильтрации путем приведения в действие вакуумного устройства к сетчатому телу 16D, которое служит вакуумной камерой, прилагается усилие вакуума через вакуумные шланги 20В, вакуумные трубы 20A, армирующие трубы 17 и первые соединительные отверстия 16Е пластины 16С, и таким образом усилие вакуума прилагается к пульпе S на фильтровальной ткани 11. Под воздействием этого усилия вакуума жидкая часть пульпы S, находящейся на фильтровальной ткани 11, отбирается через фильтровальную ткань 11 и последовательно через сетчатое тело 16D, первые соединительные отверстия 16Е на пластине 16С, армирующие трубы 17, вакуумные трубы 20А и вакуумные шланги 20В. В вакуумной камере (сетчатое тело 16D) вакуум снимают путем переключения продувочного механизма устройства вакуумирования обратно на атмосферное давление.
Вакуумные лотки 16, которые примыкают друг к другу в направлении перемещения F, соединены между собой. Как показано на фиг.1, между этими вакуумными лотками 16 на раме 10А корпуса устройства 10 установлен приводной цилиндр 18, такой как пневмоцилиндр. Приводной цилиндр 18 раздвигается и сжимается в направлении перемещения F и соединяется с вакуумным лотком 16, расположенным в самом начале линии по направлению движения F. Благодаря раздвиганию и сжатию приводного цилиндра 18 все вакуумные лотки 16 могут совместно выполнять возвратно-поступательное движение с заданной величиной шага как по направлению перемещения F, так и обратно, т.е. могут выполнять возвратно-поступательное движение вдоль этого направления перемещения F. Наряду с этим перемещением вперед и назад вакуумных лотков 16 происходит изгиб с упругой деформацией вакуумных шлангов 20В, а соединительные фитинги выполняют как одно целое возвратно-поступательное движение.
Как показано на фиг.1, между этими вакуумными лотками 16 на раме 10А корпуса устройства 10 установлен приводной цилиндр 18, такой как пневмоцилиндр. Приводной цилиндр 18 раздвигается и сжимается в направлении перемещения F и соединяется с вакуумным лотком 16, расположенным в самом начале линии по направлению движения F. Благодаря раздвиганию и сжатию приводного цилиндра 18 все вакуумные лотки 16 могут совместно выполнять возвратно-поступательное движение с заданной величиной шага как по направлению перемещения F, так и обратно, т.е. могут выполнять возвратно-поступательное движение вдоль этого направления перемещения F. Наряду с этим перемещением вперед и назад вакуумных лотков 16 происходит изгиб с упругой деформацией вакуумных шлангов 20В, а соединительные фитинги выполняют как одно целое возвратно-поступательное движение.
Как показано на фиг.1 и 3, на корпусе 10 устройства предусмотрено несколько вращающихся опорных колес 19, которые поддерживают вакуумные лотки 16, и эти опорные колеса 19 разделены в поперечном направлении относительно фильтровальной ткани 11. Опорные колеса касаются снизу пары направляющих 22, размещенных под обоими концами вакуумных лотков 16 в поперечном направлении и проходящих в направлении перемещения F. Вакуумные лотки 16 могут выполнять на этих свободно вращающихся опорных колесах 19 возвратно-поступательное движение. И наоборот, опорные колеса 19 приводятся во вращение возвратно-поступательным движением вакуумных лотков 16 под воздействием приводного цилиндра 18. Опорные колеса 19, которые все имеют одинаковую форму и одинаковые размеры, установлены таким образом, чтобы иметь возможность свободно вращаться вокруг горизонтальной оси вращения, проходящей в поперечном направлении через скобы и т.п., размещенные будучи обращенными вперед на раме 10А корпуса устройства 10. Опорное колесо 19 может иметь форму диска или форму колонны с короткой осью и отцентрировано на этой оси вращения. Опорные колеса 19 размещены так, чтобы формировать два параллельных ряда, которые тянутся в направлении перемещения F вдоль обеих кромок нижней поверхности вакуумных лотков 16 в поперечном направлении.
Опорные колеса касаются снизу пары направляющих 22, размещенных под обоими концами вакуумных лотков 16 в поперечном направлении и проходящих в направлении перемещения F. Вакуумные лотки 16 могут выполнять на этих свободно вращающихся опорных колесах 19 возвратно-поступательное движение. И наоборот, опорные колеса 19 приводятся во вращение возвратно-поступательным движением вакуумных лотков 16 под воздействием приводного цилиндра 18. Опорные колеса 19, которые все имеют одинаковую форму и одинаковые размеры, установлены таким образом, чтобы иметь возможность свободно вращаться вокруг горизонтальной оси вращения, проходящей в поперечном направлении через скобы и т.п., размещенные будучи обращенными вперед на раме 10А корпуса устройства 10. Опорное колесо 19 может иметь форму диска или форму колонны с короткой осью и отцентрировано на этой оси вращения. Опорные колеса 19 размещены так, чтобы формировать два параллельных ряда, которые тянутся в направлении перемещения F вдоль обеих кромок нижней поверхности вакуумных лотков 16 в поперечном направлении. Опорные колеса 19 в каждом ряду расположены через одинаковые интервалы в направлении перемещения F. В то же время опорные колеса 19 в каждом из рядов выровнены, образуя пары в поперечном направлении, и опорные колеса 19, которые образуют пары, имеют одинаковую высоту по вертикали и расположены на одинаковых позициях в направлении перемещения F.
Опорные колеса 19 в каждом ряду расположены через одинаковые интервалы в направлении перемещения F. В то же время опорные колеса 19 в каждом из рядов выровнены, образуя пары в поперечном направлении, и опорные колеса 19, которые образуют пары, имеют одинаковую высоту по вертикали и расположены на одинаковых позициях в направлении перемещения F.
Согласно варианту реализации устройства вакуумной фильтрации, описанному выше, предусмотрено несколько соединительных фитингов 20, и вакуумные шланги 20В каждого из соединительных фитингов 20 размещены таким образом, что они проходят в противоположные стороны относительно средней части вакуумного лотка 16 в поперечном направлении. Поэтому каждый из вакуумных шлангов 20В деформируется при возвратно-поступательном движении вакуумного лотка 16, и даже в том случае, когда восстанавливающие усилия, которые должны восстанавливать вакуумные лотки 20В, воздействуют на вакуумный лоток 16, существует возможность взаимно погасить эти силы на вакуумном лотке 16. Поэтому благодаря восстанавливающим силам вакуумных шлангов 20В можно свести к минимуму усилия, воздействующие на вакуумный лоток 16 в направлении, пересекающем направление перемещения. В то же время можно свести к минимуму момент вращения, который заставляет вакуумный лоток 16 раскачиваться вдоль поверхности плоской части 16А. То есть существует возможность ограничить вероятность возникновения ситуации, при которой возникают помехи перемещению вакуумных лотков 16, и существует возможность обеспечить прямолинейную стабильность вакуумных лотков 16.
Поэтому благодаря восстанавливающим силам вакуумных шлангов 20В можно свести к минимуму усилия, воздействующие на вакуумный лоток 16 в направлении, пересекающем направление перемещения. В то же время можно свести к минимуму момент вращения, который заставляет вакуумный лоток 16 раскачиваться вдоль поверхности плоской части 16А. То есть существует возможность ограничить вероятность возникновения ситуации, при которой возникают помехи перемещению вакуумных лотков 16, и существует возможность обеспечить прямолинейную стабильность вакуумных лотков 16.
Таким образом, в настоящем варианте реализации даже при такой конструкции, при которой корпус 10 устройства имеет пару направляющих 22, которые проходят в направлении перемещения F под обоими концами вакуумных лотков 16 в поперечном направлении, и опорные колеса 19, которые размещены на направляющих 22 и поддерживают вакуумные лотки 16, соприкасаясь с ними снизу, имеется возможность ограничить неравномерность износа, при которой величина износа направляющих 22 и опорных колес 19 различается на каждом крае в поперечном направлении. Можно также ограничить наклон вакуумных лотков 16 и можно ограничить вероятность возникновения ситуации, при которой фильтровальная ткань 11 легко движется неравномерно и создаются препятствия перемещению вакуумных лотков 16. В дополнение к этому, даже в случае когда, например, предусмотрены направляющие, которые ограничивают боковую поверхность вакуумных лотков 16 в направлении перемещения, существует возможность ограничить наклон вакуумных лотков 16, описанный выше. Таким образом, можно ограничить помехи перемещению вакуумных лотков 16, связанные с ситуацией, при которой направляющая скребет боковую поверхность вакуумных лотков 16. Кроме того, поскольку применяется множество соединительных фитингов 20, существует возможность поддерживать вакуумное усилие от вакуумного лотка 16 на пульпе S даже при небольшом диаметре вакуумных шлангов 20В этих соединительных фитингов 20. В этом случае, поскольку восстанавливающая сила вакуумных шлангов 20В становится небольшой, имеется возможность дополнительно ограничить вероятность возникновение ситуации, описанной выше, при которой создаются помехи перемещению вакуумного лотка 16, и в то же время становится возможным обеспечить экономию пространства под вакуумными лотками 16, и становится возможным добиться экономии затрат времени на техническое обслуживание этой части.
Можно также ограничить наклон вакуумных лотков 16 и можно ограничить вероятность возникновения ситуации, при которой фильтровальная ткань 11 легко движется неравномерно и создаются препятствия перемещению вакуумных лотков 16. В дополнение к этому, даже в случае когда, например, предусмотрены направляющие, которые ограничивают боковую поверхность вакуумных лотков 16 в направлении перемещения, существует возможность ограничить наклон вакуумных лотков 16, описанный выше. Таким образом, можно ограничить помехи перемещению вакуумных лотков 16, связанные с ситуацией, при которой направляющая скребет боковую поверхность вакуумных лотков 16. Кроме того, поскольку применяется множество соединительных фитингов 20, существует возможность поддерживать вакуумное усилие от вакуумного лотка 16 на пульпе S даже при небольшом диаметре вакуумных шлангов 20В этих соединительных фитингов 20. В этом случае, поскольку восстанавливающая сила вакуумных шлангов 20В становится небольшой, имеется возможность дополнительно ограничить вероятность возникновение ситуации, описанной выше, при которой создаются помехи перемещению вакуумного лотка 16, и в то же время становится возможным обеспечить экономию пространства под вакуумными лотками 16, и становится возможным добиться экономии затрат времени на техническое обслуживание этой части.
Здесь, в настоящем варианте реализации, в связи с тем что вакуумные шланги 20В расположены симметрично относительно середины вакуумного лотка 16 в поперечном направлении, восстанавливающая сила, связанная с каждым из вакуумных шлангом 20В, может быть надежно погашена, и можно значительно улучшить прямолинейную стабильность этих вакуумных лотков 16.
Кроме того, поскольку вакуумные шланги 20В соединены снизу с вакуумными лотками 16 в изогнутом состоянии, так чтобы формировать выпуклость, обращенную вниз относительно направления перемещения F фильтровальной ткани 11, вакуумные шланги 20В проходят в направлении вдоль поверхности фильтровальной ткани 11 и вакуумных лотков 16. Поэтому становится возможным гарантировать наличие пространства под вакуумными лотками и становится возможным ограничить затраты времени на техническое обслуживание этой части.
Кроме того, поскольку вакуумные шланги 20В соединены снизу с вакуумными лотками 16 таким образом, чтобы отделяться друг от друга, когда средняя часть вакуумных лотков 16 в поперечном направлении расположена между ними, вакуумные шланги 20В никогда не будут пересекаться под вакуумными шлангами 16. Поэтому становится возможным надежно избежать пересечения между каждым из вакуумных шлангов 20В, и в то же время гарантировать наличие даже большего пространства под вакуумными лотками 16.
Поэтому становится возможным надежно избежать пересечения между каждым из вакуумных шлангов 20В, и в то же время гарантировать наличие даже большего пространства под вакуумными лотками 16.
Кроме того, поскольку вакуумные лотки 16 конфигурированы таким образом, чтобы размещаться в направлении F перемещения фильтровальной ткани 11, и вакуумные шланги 20В соединены снизу с каждым из вакуумных лотков 16, уменьшая диаметр вакуумных шлангов 20В за счет применения большого количества соединительных фитингов 20, можно ограничить пересечение между соседними вакуумными шлангами 20В в направлении F перемещения, и можно уменьшить количество видов деталей, образующих это устройство вакуумной фильтрации, и можно уменьшить стоимость и снизить затраты на эксплуатацию и техническое обслуживание.
Как показано выше, настоящий вариант реализации иллюстрирует конструкцию устройства вакуумной фильтрации, в котором несколько вакуумных лотков 16 конфигурированы таким образом, чтобы размещаться в направлении F перемещения фильтровальной ткани 11, и соединительные фитинги присоединены к каждому из вакуумных лотков 16 снизу.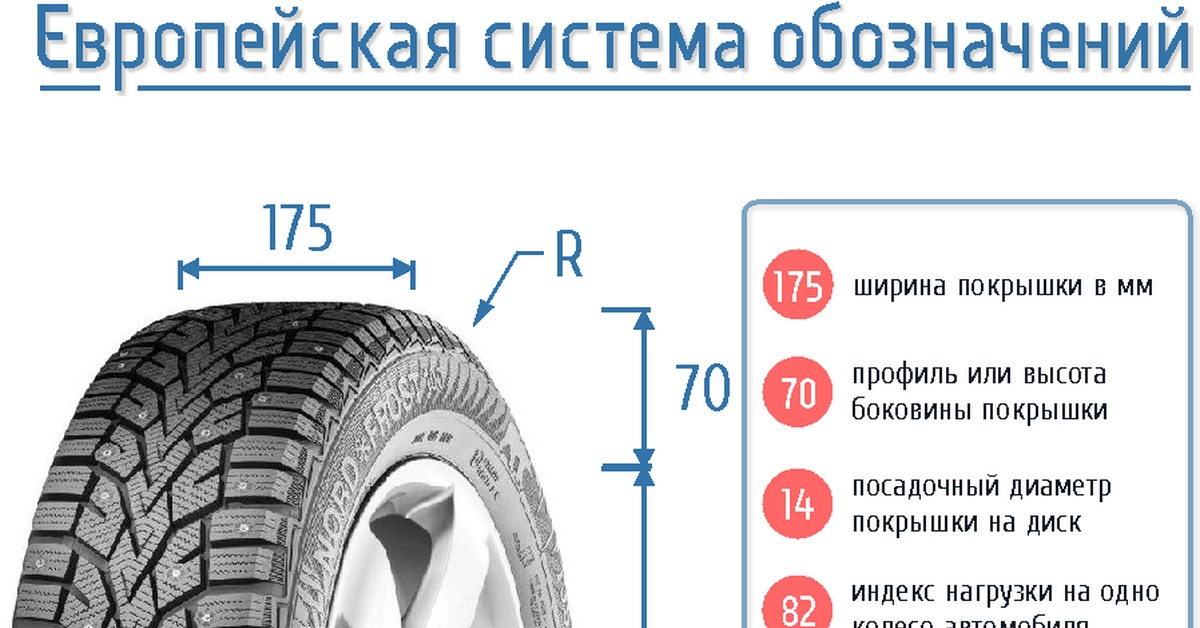 В этом случае можно ожидать возникновения проблемы, связанной с сужением пространства под вакуумными лотками 16 и, таким образом, возможного возникновения пересечений между соседними вакуумными шлангами 20В в направлении F перемещения.
В этом случае можно ожидать возникновения проблемы, связанной с сужением пространства под вакуумными лотками 16 и, таким образом, возможного возникновения пересечений между соседними вакуумными шлангами 20В в направлении F перемещения.
Однако в настоящем варианте реализации, описанном выше, поскольку диаметр вакуумных шлангов 20В может быть уменьшен за счет применения большого количества соединительных фитингов 20, существует возможность гарантировать наличие пространства под вакуумными лотками 16 и предотвратить пересечение между вакуумными шлангами 20В и можно также не допустить увеличения затрат времени на техническое обслуживание. В то же время имеется возможность уменьшить количество и типы деталей, которые образуют это устройство вакуумной фильтрации. Таким образом, существует возможность уменьшить стоимость этого устройства и снизить затраты на его эксплуатацию и техническое обслуживание.
Кроме того, благодаря армирующим трубам 17, которые прикреплены к дну вакуумных лотков 16, расположены в поперечном направлении относительно фильтровальной ткани 11 и имеют ребристую форму, возможно повышение жесткости вакуумных лотков 16, противостоящих деформации под воздействием внешних усилий. Поэтому даже в том случае, при котором восстанавливающие силы, создаваемые вакуумными шлангами 20В, воздействуют на плоскую часть 16А вакуумных лотков 16, деформируя эту плоскую часть 16А, плоской части 16А может быть придана жесткость, которая способна противостоять этим восстанавливающим силам. Поэтому существует надежная возможность не допустить помех перемещению вакуумных лотков 16, и в то же время можно ограничить вероятность неравномерного движения фильтровальной ткани 11.
Поэтому даже в том случае, при котором восстанавливающие силы, создаваемые вакуумными шлангами 20В, воздействуют на плоскую часть 16А вакуумных лотков 16, деформируя эту плоскую часть 16А, плоской части 16А может быть придана жесткость, которая способна противостоять этим восстанавливающим силам. Поэтому существует надежная возможность не допустить помех перемещению вакуумных лотков 16, и в то же время можно ограничить вероятность неравномерного движения фильтровальной ткани 11.
Кроме того, вакуумные трубы 20A соединены с нижней поверхностью пластины 16С вакуумного лотка 16 посредством армирующих труб 17, которые обладают жесткостью более высокой, чем жесткость этой пластины 16С, которая противостоит деформации. Поэтому даже в том случае, когда соединительная часть между вакуумными трубами 20A и вакуумным лотком 16 повторно подвергается нагрузке, связанной или с перемещением вакуумных лотков 16 при использовании этого устройства вакуумной фильтрации, или с восстанавливающими силами вакуумных шлангов 20В, имеется возможность ограничить вероятность поломки этой соединительной части.
В частности, в случае, при котором вакуумная труба 20A непосредственно будет соединена с пластиной 16С, которая обладает небольшой жесткостью для того чтобы противостоять деформации, вызванной внешними силами, пластина 16С легко деформируется под воздействием восстанавливающих сил вакуумных шлангов 20В и остаточных напряжений, легко накапливающихся под воздействием повторяющихся возвратно-поступательных движений вакуумных лотков 16. Таким образом, увеличивается нагрузка на соединительную часть и эту соединительную часть оказывается сравнительно легко повредить.
В отличие от этого в случае настоящего варианта реализации, когда вакуумные трубы 20А соединены с армирующими трубами 17, обладающими сравнительно большой жесткостью для противостояния деформации, вызванной внешними воздействиями, восстанавливающие силы вакуумных шлангов 20В воздействуют на армирующие трубы 17, но не воздействуют непосредственно на вакуумные лотки 16.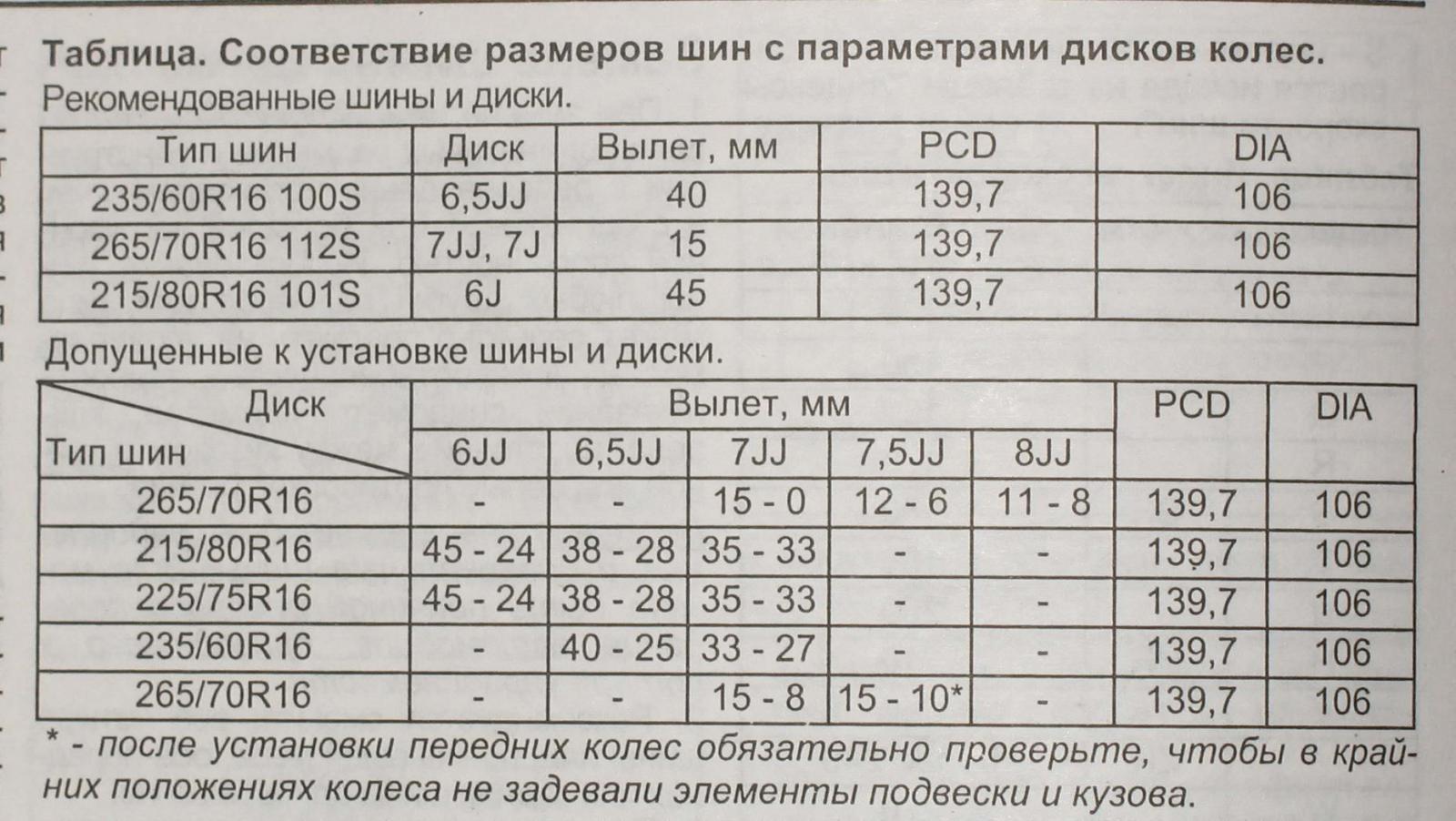 Поскольку армирующая труба 17 сама по себе обладает большой жесткостью, деформация, связанная с восстанавливающими силами или накоплением остаточных напряжений, может быть ограничена. Поэтому может быть ограничена вероятность повреждения соединительной части между вакуумными трубами 20A и нижней поверхностью пластины 16С, и поэтому можно обеспечить как увеличение срока службы устройства вакуумной фильтрации, так и уменьшение затрат времени на техническое обслуживание устройства вакуумной фильтрации.
Поскольку армирующая труба 17 сама по себе обладает большой жесткостью, деформация, связанная с восстанавливающими силами или накоплением остаточных напряжений, может быть ограничена. Поэтому может быть ограничена вероятность повреждения соединительной части между вакуумными трубами 20A и нижней поверхностью пластины 16С, и поэтому можно обеспечить как увеличение срока службы устройства вакуумной фильтрации, так и уменьшение затрат времени на техническое обслуживание устройства вакуумной фильтрации.
В случае устройства вакуумной фильтрации, имеющего большие размеры и фильтровальную ткань 11, ширина которой равна 2500 мм или больше, возрастает, конечно, количество фильтрата, отобранного из пульпы S, и требуется, чтобы усилие вакуума, приложенное к пульпе S, было в течение короткого периода времени одинаковым в поперечном направлении относительно вакуумных лотков 16. Для того чтобы добиться этого, можно рассмотреть использование конструкции, в которой применено множество вакуумных труб 20A таким образом, чтобы они были разделены промежутками в поперечном направлении на нижней поверхности пластины 16С, которая образует вакуумный лоток 16, соединительные трубы, которые соединяют вакуумные трубы 20A между собой, и один вакуумный шланг 20В, присое
Giugiaro design шины.
 Шины Vredestein
Шины VredesteinЛетняя шина Vredestein Ultrac Vorti является результатом тесного сотрудничества с известной дизайнерской компанией Giugiaro Design, которая без малого два десятка лет принимает самое активное участие в разработке новых моделей. Название этой шины было придумано итальянским дизайнерским ателье. Vorti – это производное от слова Vortex, переводимое, как «вихрь». По мнению дизайнеров, именно этим словом можно охарактеризовать способность шины безболезненно переносить любые нагрузки при движении на высокой скорости. Помимо этого, протекторный рисунок содержит элементы, формы которых имеют очень много общего с визуальным эффектом, возникающим при вращении вертолетных лопастей.
Сочетание стильного внешнего вида и функциональности
Работая над внешним обликом протекторного рисунка, итальянские дизайнеры старались придать ему спортивность и инновационность. Однако помимо эффектного дизайна асимметричный рисунок протектор содержит целый ряд оригинальных технических решений, обеспечивших шине превосходные эксплуатационные характеристики.
В частности, внешние стенки дренажных каналов имеют два угла наклона. Один из них составляет 45 градусов, что позволило повысить жесткость протекторных блоков. Результатом этого технического решения стало значительное повышение стабильности формы блоков под воздействием боковых ускорений, что благотворно сказалось на курсовой устойчивости при прохождении поворотов.
Стенки поперечных дренажных каналов также содержат дополнительный угол, составляющий 10 градусов. Таким способом голландские шинники сумели снизить сопротивление качения, а также уровень шума. Кроме того, такая форма стенок поперечных каналов позволила значительно улучшить распределение внешнего давления по пятну контакта, в результате чего повысилась стойкость протектора к неравномерному износу.
Еще одной отличительной особенностью асимметричного рисунка данной модели являются стреловидные канавки, расположенные на центральном продольном ребре. Эти протекторные элементы позволили существенно улучшить курсовую устойчивость при движении на высокой скорости.
Два варианта – для передней и задней оси
Данная модель предназначена для мощных спортивных легковых автомобилей, на многие из которых устанавливаются шины разных типоразмеров. Как правило, на заднюю ось ставятся более широкие шины, что позволяют эффективно справляться с высокой мощностью двигателя. Специально для таких автомобилей голландский производитель предлагает два варианта шины Vredestein Ultrac Vorti , которые имеют значительные различия, как по рисунку протектора, так и по конструкции. Шины, предназначенные для передней оси, отличаются повышенными дренажными свойствами. Иначе говоря, они удаляют большую часть воды с поверхности дорожного полотна. В свою очередь, шины, устанавливаемые на заднюю ось, обеспечивают максимальное сцепление с дорогой. Именно поэтому их рисунок обеспечивает больший, чем у передних шин, коэффициент пустотности. Кроме того, у шин для задней оси несколько изменен угол расположения брекерного слоя. Это позволило еще больше увеличить размеры пятна контакта.
4-радиусная форма профиля
Одной из ключевых особенностей этой модели является многорадиусный профиль. У большинства легковых шин контур включает в себя три радиуса. Однако профиль этой модели содержит четыре радиуса, что позволило значительно увеличить размеры пятна контакта, а также улучшить равномерность распределения внешней нагрузки по нему.
Основные достоинства этого технического решения проявляются при прохождении поворотов, когда основная нагрузка приходится на плечевые зоны протектора, что приводит к изменению, как формы, так и размеров пятна контакта. Новая форма профиля позволила не только уменьшить степень этих изменений, но и сделать этот процесс более плавным. Результатом таких технических ухищрений стало стабильное и прогнозируемое поведение при прохождении поворотов.
Основные особенности шины Vredestein Ultrac Vorti
— Асимметричный рисунок протектора, разработанный с участием итальянских дизайнеров, отличается превосходной эффективностью в любых дорожных условиях;
— Различные варианты для передней и задней оси позволяют обеспечить идеальный баланс эксплуатационных характеристик;
— Резиновая смесь с повышенным содержанием кремнийсодержащих элементов, благодаря чему шина демонстрирует отличное сцепление с мокрой поверхностью при повышенной стойкости к абразивному износу;
— Очень широкие продольные дренажные канавки обеспечивают быстрый отвод воды из пятна контакта, обеспечивая тем самым отличную стойкость к аквапланированию даже на высокой скорости;
— 4-радиусная форма профиля шины обеспечивает высокую стабильность и прогнозируемость поведения, особенно при прохождении поворотов.
* Внимание: летние шины не российского происхождения могут быть промаркированы обозначением M+S
Возможно, вас заинтересуют также следующие модели.
Перед началом сезона Vredestein порадовал наших журналистов презентацией сразу трех новинок. Две премьеры российские, одна – мировая.
Под ласковым андалузским солнцем нам предложили попробовать в деле скоростные шины Sportrac третьего поколения, уже хорошо известные в Европе, Ultrac Sessanta класса UHP, выпуск которых был приурочен к прошлогоднему юбилею фирмы, и абсолютно новую их модификацию Ultrac SUV Sessanta для городских внедорожников.
Сначала попробуем
В отличие от некоторых своих более именитых коллег, голландцы доверились нашему водительскому мастерству. Расписаться в договоре, согласно которому ответственность за возможные происшествия мы принимаем на себя (общепринятая практика), нас вежливо попросили, а затем отпустили в «свободное плавание». С берега Средиземного моря мы самостоятельно отправились в глубь Испании на скоростную трассу Ascari Race Resort. Ни на платной магистрали, ни на горном серпантине шины о себе не напомнили. Что Land Rover, что Mercedes-Benz M-class «просто ехали», подчиняясь действиям водителя. Вывод, сделанный «по дороге»: на асфальте шины на удивление тихи и комфортны.
Зато на треке, точнее на трех разных его участках и прилегающей площадке, досталось им по полной программе.
Начали с «мокрого» теста: змейка, полукруг и коридор с торможением на залитом водой асфальте. Единственное замечание к BMW на шинах Ultrac Sessanta: мало дали покататься. На «закрученной» дистанции аквапланирование было просто не ощутить. А вот паркетники в одном вираже немного сносило. Винить в этом Ultrac SUV Sessanta грешно: у машин такого класса и вес побольше, и центр тяжести выше.
Перебираемся на первое скоростное кольцо. Следуя указаниям инструкторов, перед поворотами притормаживаем, газ прибавляем на выходе, приближаем траекторию к оптимальной. Если ехать по правилам, вопросов не возникает. А если нет? Ведь достоинство шины в даваемом ею шансе исправить допущенную ошибку.
Характер у Sessanta оказался «мягким». Силовое вкатывание в поворот не приводит к резкому срыву колес, машина начинает прогнозируемо «плыть». Трудно заочно сравнивать с конкурентами, но ведет себя «юбилейная» шина вполне достойно.
Пересаживаемся на полноприводники. Второе кольцо имеет большие перепады высот и один очень неприятный левый поворот. Машины не для кольцевых гонок, стиль вождения невольно выбираешь соответствующий. Audi Q7 раздражает алгоритмом своей автоматической коробки, ML куда более спортивен. Немного напугал Land Rover. Тяжелая машина не приспособлена для трека, от резкого торможения тормоза начинают перегреваться и «шалить». Косвенное подтверждение качества шин: слабым местом оказываются не они, а колодки.
Катание на суперкаре с инструктором за рулем мне пришлось пропустить: если бы пустили за руль, может, и постарался бы втиснуть свое непомерное тело в мини- атюрный салон, а сидеть калачиком справа – удовольствие не для меня.
«Практические занятия» показали, что звание Ultra High Performance шины Vredestein носят по праву. Лучше или хуже конкурентов, не скажу: нужны более длительные тесты и точные измерения. По ощущениям — сопоставимы.
От дел к слову
Нарушая сложившуюся традицию, техническую презентацию шин провели после серии заездов. Итак, знакомимся.
Как и все последние разработки Vredestein, семейство Ultrac Sessanta создавалось при непосредственном участии итальянского ателье Giugiaro Design. Конструкторы шины определяли основные внешние особенности рисунка протектора (приблизительное соотношение канавок и блоков, их ориентировочное сечение и т.п.). Дизайнеры придавали задумкам художественное совершенство. Затем конструкторы выбирали наиболее пригодные с их точки зрения варианты и «учили их ходить». Вопреки складывающейся в отрасли традиции, шины UHP от Vredestein внешне не только не скучны, но динамичны и агрессивны.
Основное внимание при создании Ultrac Sessanta уделялось управляемости на сухой и мокрой дороге. В основу конструкции заложены две идеи: резиновая смесь протектора собственной разработки, «настраиваемая на управляемость» (HTTC – Handling Tuned Tread Compound), и «усложненный продольный рисунок протектора (SLD – Sophisticated Longitudinal Design).
Смесь НТТС содержит силику и разные виды активированного углерода, обеспечивающие отличную управляемость как на сухой, так и на мокрой дороге. За счет большой активной поверхности частицы углерода хорошо связываются с полимерами, образуя очень прочный материал протектора. Благодаря НТТС шины Ultrac Sessanta способны в повороте передавать экстремально большую силу, величина которой в общем случае пропорциональна максимально возможной скорости прохождения поворота. Особенность новых шин – в плавности нарастания силы по мере роста угла бокового увода и, соответственно, в предсказуемости поведения. Более того, сила, удерживающая шину в повороте, не снижается при больших значениях увода, а остается на достаточно высоком уровне. За счет этого появляется возможность дополнительной точной корректировки курса движения. Свою лепту в такое улучшение характеристик, помимо смеси, вносит и рисунок протектора, и внутреннее устройство шины.
Рисунок протектора Ultrac Sessanta – асимметричный ненаправленный: центральную продольную канавку дополняет множество изогнутых к наружному плечу диагональных «перьев». Чем-то похоже на расправленное крыло большой птицы. В пятне контакта «косые канавки» никогда не оказываются замкнутыми, так как их длина больше длины «отпечатка» шины на дороге. Таким образом, рисунок можно рассматривать как имеющий четыре продольных канавки. Отличное решение для борьбы с аквапланированием. На мой вопрос о разной из-за различного поперечного сечения канавок их пропускной способности специалисты компании ответили, что даже минимального сечения достаточно. А разница не вызовет «всплывания» одного из колес. Но если это так, то излишняя ширина красиво расширяющейся канавки – это меньшая площадь «позитива», большие удельные нагрузки, следовательно, и больший износ. Возможно, этим ростом можно пренебречь из-за его незначительности. Тем более что срок службы – далеко не самый важный показатель шин Ultra High Performance.
А вот другое (чисто умозрительное) заключение хотелось бы проверить. На пятнах контакта хорошо видно, что «косые» канавки смотрят в одну сторону. На ровном асфальте это не имеет никакого практического значения.
Вспомните иные наши дороги и наши методы их ремонта. Продольная «насечка» на асфальте, остающаяся от проезда гусеничной техники или снятия верхнего слоя старого асфальта, работая, как рельсы, теоретически может «сдвинуть» автомобиль в сторону. Надеюсь, что я ошибаюсь, но тем, кто приобретет Ultrac Sessanta, посоветую поначалу присмотреться к поведению шин в этих необычных и, к счастью, редких дорожных условиях. С другой стороны, оригинальный рисунок имеет массивное наружное плечо. Позитива (резины) в нем на четверть больше, чем во внутреннем. А именно наружное плечо наиболее загружено в повороте. Отсюда лучшая курсовая устойчивость и управляемость на кривых.
Среди других технологических решений стоит отметить уже ставшую традиционной для голландцев конструкцию боковины HTS+ (Handled Tuned Sidewall – настроенная на управляемость боковина). За счет двухслойного текстильного каркаса боковая часть шины сделана более жесткой, что позволяет ей лучше противостоять нагрузкам, возникающим при резких поворотах. Технология была впервые опробована на шинах Sportrac 2 и в последующих моделях дважды усовершенствована. HTS+ обеспечивает минимальную деформацию боковины и максимальное сопротивление поперечным силам за счет большей осевой жесткости. Как следствие, улучшается управляемость в критических ситуациях.
Нельзя не упомянуть и другую конструктивную особенность – оболочку из металлокорда. Она отвечает за безопасность, в частности, за точность рулевого управления. Брекер должен быть максимально жестким, каркас – максимально приближенным к протектору. Для достижения первой цели между слоями корда используется специальный состав смеси и подбирается оптимальный угол наклона нитей, для второй – резина наносится только с одной стороны наружного нейлонового слоя. Уменьшение количества резины между слоями приближает каркас в целом к поверхности дороги.
Шина Ultrac Sessanta появилась сражу же в широком диапазоне типоразмеров. Это стало возможным за счет использования трехмерной (3D CAD) системы компьютерного моделирования P3DD (Parametrical 3Dimensions Design – параметрическое трехмерное конструирование). В сочетании с обширной базой данных система позволяет быстро реагировать на потребности рынка, автоматически оптимизировать любой необходимый размер новой шины. С помощью метода конечных элементов (FEM – Finite Elements Method) вычисляется и оптимальное пятно контакта.
Именно набор средств компьютерного обеспечения позволил в кратчайшие сроки дополнить линейку Ultrac Sessanta семейством шин для спортивно-утилитарных автомобилей – Ultrac SUV Sessanta.
По сравнению со спортивными автомобилями, очень модные ныне паркетники и полноценные внедорожники предъявляют к шинам более высокие требования. Ведь зачастую двигатель, достойный суперкара, устанавливается на куда более тяжелый автомобиль. В целом, аналогичные шинам Ultrac Sessanta новички 2007 года отличаются большей шириной и повышенной грузоподъемностью. Пока предложено 12 типоразмеров Ultrac SUV Sessanta (от 225/65ZR17 108 V XL до 295/30ZR22 103 Y XL). «Легковых» типоразмеров больше – 36 (от 225/45ZR17 94 Y XL до 295/25ZR20 95 Y XL).
«Целевая аудитория» шин Sportrac 3 более демократична. Предназначаются они более скромным, но достаточно скоростным автомобилям: Alfa Romeo 159, Volvo S40 и V50, VW Passat и их одноклассникам.
Упор при разработке делался на улучшение сцепных характеристик на мокрой дороге. Рисунок протектора (V-образные канавки в центре и мощные плечевые зоны) также разрабатывался итальянцами. Шину отличает равномерное распределение давления в пятне контакта, обеспечивающее хорошие сцепные качества, снижение износа и шума. В состав резиновой смеси включена силика (high-silica solution V). С боковинами HTS мы уже познакомились. 16-дюймовые шины снабжены кольцевым ребром, защищающим обод колеса от повреждений о бордюрный камень. Шина характеризуется хорошими сцепными свойствами, надежной управляемостью в поворотах. Sportrac 3 – третье поколение семейства, дебютировавшего в 1998 году. Именно с него началось сотрудничество Vredestein с Giugiaro Design. В 2003 году линейка была обновлена и ныне соответствует всем современным требованиям. В России в этом году будет продаваться всего четыре типоразмера Sportrac 3: 205/50R16 87V, 205/55R16 91V, 205/60R16 92V и 215/55R16 XL 97V.
Vredestein Bonden
История торговой марки Vredestein заслуживает уважения. Началась она после Второй мировой войны: в июне 1946 года был основан голландско-американский завод Vredestein по производству шин для легковых автомобилей. Его крупнейшими акционерами стали голландская компания N.V. Rubberfabriek Vredestein и американская (и тогда самостоятельная) BF Goodrich. Многое изменилось за шесть десятков лет. Небольшая, по мировым меркам, компания практически весь этот срок умудрялась противостоять транснациональным гигантам. Выстоять в труднейшей конкурентной борьбе помогали и традиционное голландское качество, и умелая маркетинговая политика, и собственные разработки. Большинство владельцев Porsche три с лишним десятилетия знают «запаску-гармошку» от Vredestein: колесо размером с «докатку» в походном положении, при накачивании превращающееся в полноразмерное.
В начале 90-х голландцы разработали первую низкопрофильную «дождевую» шину (Aquatrac) с индексом скорости Т (до 190 км/ч). Удивляет Vredestein и дизайном рисунка протектора – сотрудничество с всемирно известным ателье Giugiaro Design продолжается уже более 10 лет.
Помимо легковых, Vredestein выпускает сельскохозяйственные и мотоциклетные шины. В 2005 году «гордой независимости» компании, к сожалению (а может, к счастью), пришел конец: она влилась в российский Amtel. «Амтел-Фредештайн» Н.В.» занял одно из лидирующих мест среди крупнейших европейских производителей, приблизившись к ведущей мировой десятке. Вполне вероятно, что голландские технологии, помноженные на российские деньги, помогут новому объединению укрепить свои позиции на мировом рынке.
Race Resort Ascari
В сентябре 2003 года на юге Испании, неподалеку от исторической Ронды и совсем рядом с Марбеллой открылся первый в мире гоночный курорт «Аскари», объединивший в живописнейшей горной долине современную трассу для кольцевых гонок и первоклассный гостиничный комплекс.
Кольцо построено в строгом соответствии с требованиями FIA. Оно длиннее, чем большинство трасс «Формулы 1», и имеет вдвое большее количество поворотов. Конфигурацию дистанции можно легко изменить. В день презентации трасса была разбита на три коротких отрезка. Каждый из 26 поворотов имеет свою комбинацию радиуса, подъема/спуска и поперечного уклона. Прототипами для многих из них стали такие всемирно известные «кривые», как Eau Rouge в Спа-Франкоршам, Daytona Parabola, Cops в Сильверстоуне или Druids в Оултен Парке.
Пит-лейн несколько короче – всего 8 полноценных боксов. Зато вдобавок к шоссейной имеется шестикилометровая грунтовая трасса для внедорожников и кольцо для кросса на багги.
Судить о достоинствах андалузского Cortijo не берусь. Кулинарные рецепты — из той же области. Это дело специалистов, которые, кстати, уже отметили курорт многочисленными дипломами и сертификатами.
Race Resort Ascari – это не только трассы и отель. Ascari Club, эксклюзивный автомобильный клуб с членством не более 500 человек, привлекает состоятельных автовладельцев не только возможностью отдохнуть в сказочном по красоте месте после получения очередной дозы адреналина. Члены клуба имеют целый список «автоспортивных привилегий», включая хранение собственных гоночных автомобилей. К их услугам лучшие номера и все возможности курорта. Плюс новые бизнес-контакты. Куда демократичнее Ascari Race Academy – гоночная академия, в которой можно овладеть навыками управления разными типами автомобилей, такими, как BMW 3-series, Lotus Elise, Radical SR3, Formula Reynard, Ascari KZ1 и Benetton F1. В программах обучения учитывают время, которым располагает клиент, уровень его подготовки и, разумеется, его потребности и пожелания.
Если же вы пожелаете провести на курорте спортивный праздник, презентацию или, чем черт не шутит, международную гонку, вам на помощь придет служба обеспечения мероприятий Ascari Event Services. Она же забронирует места в ближайших гостиницах, организует перевозку людей и техники, устроит катание на воздушных шарах, экскурсии в римский амфитеатр или пещеры с доисторической наскальной живописью. Или за 15 минут доставит вас на средиземноморское побережье.
Первые мысли по приезде на трек: не назван ли он в честь легендарного Альберто Аскари? Или, может, он принадлежит его потомкам? Или, что маловероятно, великий гонщик сам из этих мест?
Организаторы презентации затруднились с ответами на мои вопросы. Выручили «местные». Справедливым оказалось только первое предположение. Причем с оговоркой.
В 1994 году в Англии была основана компания Ascari Cars, действительно названная в честь Альберто Аскари. Группа гоночных инженеров взялась за создание суперкара. Первую ограниченную серию в 20 Ascari Ecosse выпустили в 1998 году. В 1999-м была создана команда Ascari, позднее участвовавшая в гонках спортпрототипов, включая чемпионат FIA, суточные гонки в Ле-Мане и Дайтоне, серии EuroBoss разных лет. В 2000 году Ascari Cars построила новый завод в английском Бенбери для выпуска новой модели суперкара Ascari KZ1. Кроме того, основатель фирмы голландский инженер Клаас Зварт приобрел в Андалусии 174 га земли и построил собственную гоночную трассу, на которой желающие могут в полной мере оценить достоинства спроектированного и построенного им самим болида.
Так что, перефразируя советского классика, Rase Resort посвящается «товарищу Аскари – человеку, автомобилю и заводу».
Характеристики Ascari Race Circuit
Длина, м – 5425 Прямые, м – 3133 Повороты, м – 2292 Ширина, м – 12,2 Самая длинная прямая, м – 470 Число поворотов – 26 (13 левых, 13 правых) Мин. радиус поворота, м – 7 Макс. радиус поворота, м – 899 Макс. подъем, % – 10,5 Макс. спуск, % – 11,5 Макс. поперечный уклон, % – 17,6 Мин. поперечный уклон, % – 1,0 Трасса может быть разбита на три участка – 2592 + 1341 + 1527 м
Mustang by Giugiaro
А вот еще один пример итало-нидерландского сотрудничества. Концептуальный Mustang by Giugiaro был впервые показан широкой публике в ноябре 2006 года на автошоу в Лос-Анджелесе. 500-сильный мотор подготовил Ford Racing, тормоза – Brembo, бортовую электронику – Clarion, прозрачную, но не пропускающую ультрафиолет крышу – Solutia, колеса – OZ. А шины футуристического вида – серийные. От Vredestein.
Vredestein Sportrac 3
Sportrac 3 – третье поколение зародившегося в 1998 году семейства. Динамичный рисунок протектора этих шин (созданный все теми же именитыми итальянцами) уже хорошо известен в Европе. поставки в Россию начались лишь недавно. Пока предлагается только четыре типоразмера. Все – на 16 дюймов.
Vredestein и Giugairo в этом году будут праздновать 15 лет успешного сотрудничества, за которые они успели вместе создать такие шины, как Vredestein Ultrac Sessanta и Vredestein Ultrac Vorti. В честь этого юбилея презентацию нового совместного творения было решено провести в штаб-квартире итальянской дизайн-студии в Монкальери, пригороде Турина. Всего на презентации побывало более 450 дилеров и ключевых партнеров Vredestein, а новинка получила название Vredestein Wintrac Xtreme S.
Новая модель пришла на смену шинам Vredestein Wintrac Xtreme, и по этой причине оптимизирована для того, чтобы справляться с самыми сложными условиями и экстремально низкими температурами. Специальные взаимосмыкающиеся ламели (Internal Sipe Locking Technology, ISLT) позволили еще больше повысить характеристики шин, которые сохраняют устойчивость даже на очень высоких скоростях – их индекс скорости Y говорит о том, что их можно использовать на скорости до 300 км/ч.
Как отмечает производитель, ключевые особенности новых шин – это прекрасные сцепление и управляемость на снегу, льду и слякоти. Тяговое усилие было повышено за счет увеличения числа ламелей, причем Vredestein Wintrac Xtreme S обладают ламелями двух типов. В центральных блоках они прямые, что максимизирует тягу и эффективность торможения на снегу и льду, а в плечевых зонах – зигзагообразные, что улучшает маневренность на скользких поверхностях. Рисунок протектора включает в себя две ультраширокие продольные канавки, которые в сочетании с V-образными каналами в центральной части с низким коэффициентом пустотности обечпечивают эффективный дренаж воды и высокое сцепление на мокром покрытии. Помимо маркировки M+S новые шины также отмечены символом снежинки, позволяющим продавать их как специализированные зимние покрышки во всех регионах мира.
Компаунд включает в себя улучшенную смесь полимеров, а также силан нового поколения, что позволило существенно увеличить эксплуатационный ресурс. Кроме того, эта комбинация, по словам разработчиков, помогла значительно сократить сопротивление качению.
Помимо Giugiaro, разработавшей симметричный направленный дизайн, компания Vredestein работала над новыми шинами в сотрудничестве с другими известными партнерами из автомобильной индустрии, в том числе с такими тюнинг-ателье, как Carlsson, Hamann и Mansory. Vredestein уже давно и активно сотрудничает с европейскими специалистами тюнинга, что, как говорят в компании, позволяет новым шинам, в том числе зимним, с легкостью отвечать требованиям сегмента Ultra High Performance.
Vredestein Wintrac Xtreme S будут доступны в 12 типоразмерах от 205/55 R16 до 275/30 R20.
Все 31 33 155 165 175 185 195 205 215 225 235 245 255 265 275 285 295 305 315 325 335
Все 10.5 12.5 25 30 35 40 45 50 55 60 65 70 75 80 85 100
Все R16 13 14 14С 15 15С 16 16С 17 18 19 20 21 22 24
Все Зима Лето
Все Antares Apollo Barum BFGoodrich BlackLion Bridgestone Continental Contyre Cooper Cordiant Diamondback Dunlop Ecovision Eldorado Falken Firestone Forward General Gislaved Goodride Goodyear Hankook Headway Hercules Jinyu Joyroad Karmoran Kormoran Marangoni Matador Maxxis Michelin Minerva MRF Nankang Nexen Nitto Nokian Pirelli Powertrac Premiorri Rosava Rovelo Sailun Starmaxx Tigar Toyo Triangle Tunga Viatti Vredestein Yokohama Zeta
Все Notice on line 114 Warning /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 114
Все Notice : Undefined variable: disc in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 129 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 129
Все Notice : Undefined variable: disc in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 144 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 144
Все Notice : Undefined variable: disc in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 161 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 161
Все Notice : Undefined variable: disc in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 176 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 176
Все Notice : Undefined variable: disc in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 191 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 191
Выберите — Notice /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 219 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 219
/var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 232 disabled=”disabled”> Notice : Undefined variable: auto in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 233 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 233
Notice: Undefined variable: auto in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 248 disabled=”disabled”> Notice : Undefined variable: auto in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 249 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 249
Notice: Undefined variable: auto in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 262 disabled=”disabled”> Notice : Undefined variable: auto in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 263 Warning : Invalid argument supplied for foreach() in /var/www/u0324292/public_html/сайт/catalog/view/theme/default/template/module/search_auto_tabs.tpl on line 263
Designed To Protect You (спроектировано для вашей защиты)
С 1997 года фирма Vredestein тесно сотрудничает с широко известным во всем мире итальянским дизайнерским ателье Giugiaro Design. Сам Джурджаро и его сын и преемник Фабрицио Джурджаро оказали огромное влияние как на товарный знак, так и на бренды от Vredestein. Это уникальный союз двух брендов приносит успех обеим сторонам. Наличие ассоциации между маркой Vredestein и маркой Giugiaro также способствовало тому, что марка Vredestein стала премиум брендом с такими важными характеристиками, как безопасность и дизайн.
Дизайнерское ателье Giugiaro Design отвечает за дизайн протектора и боковой части шины Vredestein Ultra High Performance. Разумеется, специалисты-инженеры фирмы Vredestein имеют решающий голос: основное предназначение шины – это обеспечение безопасности, однако шина от Vredestein сочетает в себе как безопасность, так и дизайн. Более того, результаты международных испытаний, проведенных с шинами, продемонстрировали, что привлекательность шины заключается не только в её броском внешнем облике, но и в её превосходных эксплутационных качествах, что позволило ей каждый раз занимать высокое место по результатам испытаний.
В 2012 году исполнилось 15 лет сотрудничеству компании Vredestein и Giugiaro Design. В честь этого уникального события компания Vredestein представила два новых типа летних шин, разработанных совместно с Giugiaro: Ultrac Vorti и Sportrac 5.
Сегодня знаменитая компания Vredestein занимается разработкой и изготовлением высококачественных шин для легковых и грузовых автомобилей, промышленной и сельскохозяйственной техники. Но приобрести изделия премиум-класса для многих водителей не представляется возможным. Мы же предлагаем купить бу шины Vredestein по доступной стоимости.
Особенности бренда
Благодаря постоянным внедрениям передовых технологий и постоянному совершенствованию продукции производителю удалось достигнуть невероятных результатов. Автопокрышки Фредештайн адаптированы для функционирования в самых сложных дорожных и климатических условиях. Среди других достоинств:
- мощные тормозные и сцепные характеристики;
- прекрасная управляемость транспортным средством;
- высокие показатели прочности, надежности и износостойкости;
- акустический комфорт.
Виды резины Vredestein
Значительно продлить срок службы летней резины Vredestein производителю удалось применением зигзагообразных рисунков протектора. Феноменальную грузоподъемность гарантируют специальные блоки с усиленной жесткостью. Также они надежно защищают изделие от различных механических повреждений и деформаций. А благодаря снижению шумности и вибраций езда стала особенно комфортной и приятной.
Наличие широких канавок позволяет передвигаться не только по асфальтированным дорогам, но и бездорожью.
Шины бу Vredestein для зимы гарантируют отличную проходимость и управляемость автомобилем на заснеженных трасах. Можно также рассчитывать на прекрасные сцепные свойства, большие ходовые ресурсы и качественное торможение. А благодаря легкости качения удалось уменьшить потребление топлива до 15%.
У нас можно шины Vredestein бу купить на взаимовыгодных условиях. Мы гарантируем оригинальность и высокое качество продукции. С нами всегда приятно сотрудничать.
ПОЧЕМУ ДИАМЕТР РАСПРЕДЕЛИТЕЛЬНОЙ ШЕСТЕРНИ КОЛЕНЧАТОГО ВАЛА МЕНЬШЕ ШЕСТЕРНИ РАСПРЕДВАЛА… Распределительные шестерни
Шестерню распределительного вала изготавливают из стали, чугуна или текстолита. Повернуть коленчатый вал на пол-оборота, отрегулировать зазоры для второго цилиндра.3. Поверхности шеек распределительного вала, кулачков, эксцентрика и зубьев шестерни привода масляного насоса закалены до высокой твердости.
Между шестерней и передней шейкой вала установлены распорное кольцо 3 и ограничивающий осевое перемещение вала упорный фланец 2, который привертывают болтами к передней стенке блок-картера. Распределительные шестерни большинства двигателей расположены в передней их части в специальном картере. Они необходимы для передачи вращения от коленчатого распределительному валу, валу топливного насоса, масляному насосу и другим механизмам.
Фазы газораспределения и порядок работы цилиндров
Для того чтобы действие клапанов и подача топлива соответствовали определенному положению поршня в цилиндре, зубья указанных шестерен при сборке соединяют по меткам. В некоторых двигателях вал топливного насоса и распределительный вал вращаются в противоположные стороны.
Распределительный вал представляет собой стержень с опорными шейками и кулачками, предназначенными для своевременного открытия клапанов. Чтобы облегчить установку вала в подшипники, опорные шейки его делают иногда неодинаковыми.
Кулачковые валы двигателей с верхним расположением выполняются с осевым сверлением, через которое масло подводится к опорным шейкам и кулачкам. Распределительные валы куют из обычных или легированных среднеуглеродистых сталей (марок 40, 45, 40Х и др.), или же отливают из специального чугуна. Опорные шейки, кулачки и эксцентрики закаливают аналогично шейкам коленчатых валов с применением токов высокой частоты. Осевое перемещение кулачковых валов допускается в пределахвсего 0,1—0,2 мм. Фиксацию их осуществляют специальными ограничительными фланцами или упорными болтами.
Распределительный вал изготавливают цельным или составным (для упрощения изготовления и монтажа) и устанавливают на разъемных подшипниках. Для предотвращения осевого перемещения вал обычно фиксируют в опорно-упорном подшипнике. Кулачные шайбы в ВОД обычно отковывают заодно с распределительным валом, а в СОД и МОД выполняют съемными (неразъемными или разъемными). Цепной привод (рис. 11.7, б) используют при большом расстоянии между осями коленчатого и распределительного валов, когда шестеренный привод получился бы громоздким и дорогим.
На коленчатом валу находится стальная шестерня с 28 зубьями, а на распределительном валу – текстолитовая шестерня с 56 зубьями. Pаспределительный вал вращается в 2 раза медленнее коленчатого. От осевых перемещений распределительный вал удерживается упорным стальным фланцем. По этому сверлению и пазу масло подводится из канала в головке в полость оси коромысел. Для этого надо, проворачивая коленчатый вал пусковой рукояткой, совместить метку на ободе шкива коленчатого вала с указателем на крышке распределительных шестерен.
При притирке клапанов следует очистить от отложений полость оси коромысел, каналы в четвертой стойке оси и в головке цилиндров. Такое расположение распределительного вала называется верхним, оно позволяет упростить блок цилиндров и уменьшить шум при работе механизма газораспределения. При верхнем расположении распределительный вал приводится цепью или зубчатым ремнем.
Для этого на коленчатом и распределительном валах (рис. б) установлены шкивы 14 и 16 с наружными зубьями специального профиля. Весь привод распределительного вала не нуждается в смазке; от пыли и грязи защищен легкими пластмассовыми крышками.
Зубья у шестерен косые, что вызывает появление силы, стремящейся переместить распределительный вал в осевом направлении. Коромысло изготовляют из стали или чугуна. Правильность установки механизма ВМТ газораспределения определяется зацеплением распределительных шестерен с имеющимися на них метками. Распределительные валы газораспределительных механизмов в ремонт могут поступать со следующими дефектами: износ шеек под подшипники, прогиб, износ шестерни привода масляного насоса и кулачков.
Механизм газораспределения
Шестерни изготовлены из стали. Для уменьшения шума они выполнены косозубыми. Распределительные шестерни из стали марки 45 со спиральными зубьями помещены в кожухе. Такие биметаллические подшипники часто делают свертными из стале-баббитовой ленты (ЗМЗ-53, ЗМЗ-66 и другие двигатели) и запрессовывают в гнезда, расточенные в перегородках картера.
Для некоторых двигателей распределительные валы изготовляют из чугуна, в этом случае их кулачки и шейки подвергают отбеливанию. В том случае, если распределительный и коленчатый валы вращаются в разные стороны, то промежуточная шестерня между ними отсутствует.
Что еще посмотреть:
- Раздел «Промышленная биотехнология»В качестве продуцентов используют штаммы Candida scotti и C.tropicalis. В современных биотехнологических процессах, основанных на использовании микроорганизмов, продуцентами белка служат […]
- Администрация города УльяновскаОрганы территориального общественного самоуправления входят в систему органов местного самоуправления и, следовательно, являются органами, через которые осуществляется власть народа. Одной […]
- Опасная встреча1.2 Квест «Шахтеры»: >Q. Как «по-тихому» убить Педро Нуньеса? 1.2 Квест «Механик»: >Q. Где берется квест с механиком? 1.3 Квест «Разработчики»: >Q. Где искать производителя наркотиков? 1.2 […]
F-15 против F-16 Fighting Falcon – разница и сравнение
F-15 – двухмоторный истребитель, способный развивать чрезвычайно высокие скорости и высоты, а F-16 – менее мощный, но более маневренный одномоторный истребитель.
F-15 считается одним из самых успешных и грозных истребителей, когда-либо построенных, с более чем 100 победами в воздушных боях и нулевыми потерями в воздушных боях.Он может работать в любых погодных условиях, нести различные виды оружия класса «воздух-воздух» и «воздух-поверхность» и вооружен пушкой M61 Vulcan.
F-16 – более дешевый, легкий и немного менее мощный самолет, но он был разработан с упором на простоту обслуживания и маневренность. Первоначально задуманный как истребитель дня превосходства в воздухе, он стал универсальным всепогодным многоцелевым самолетом и имеет ту же пушку M61 Vulcan, что и F-15.
Таблица сравнения
Различия – Сходства –
| F-15 | F-16 Fighting Falcon | |
|---|---|---|
|
| |
| Роль | Истребитель превосходства в воздухе | Многоцелевой истребитель |
| Производитель | McDonnell Douglas Boeing Defense, Space and Security | General Dynamics, Lockheed Martin |
| Статус | Используется в основном в США, Японии, Саудовской Аравии и Израиле | В эксплуатации |
| Национальное происхождение | США | США |
| Количество построенных | 1,198 | 4,500+ |
| Стоимость единицы | 28-30 млн долларов США | F-16A / B: 14 долларов США.6 миллионов долларов (1998 долларов), F-16C / D: 18,8 миллиона долларов (1998 долларов) |
| Также известен как | McDonnell Douglas F-15 Eagle | – |
| Максимальная скорость | Мах 2,5 | Маха 2,02 |
| Количество двигателей | 2 | 1 |
| Представлен | Январь 1976 г. | Август 1978 г. |
| Длина | 63 фута 9 дюймов | 49 футов 5 дюймов |
| Действующий номер | 222 в США; более 1000 по всему миру. | 2000+ в | США.
| Размах крыла | 42 фута 10 дюймов | 32 фута 8 дюймов |
| Снаряженная масса | 20200 кг | 12000 кг |
| Вооружение | 20-мм 6-ствольная пушка Гатлинга M61 Vulcan, бомбы, ракеты, десантные баки. | 20-мм (0,787 дюйма) 6-ствольная пушка Гатлинга M61 Vulcan, реактивные снаряды, снаряды, бомбы |
| Скороподъемность | 50, 000+ фут / мин | 50000 фут / мин |
| Боевой радиус | 1222 мили | 340 миль |
| Практический потолок | 65000 футов | 50,000+ футов |
| Первый полет | 27 июля 1972 г. | 20 января 1974 года |
| Введение (из Википедии) | McDonnell Douglas F-15 Eagle – американский двухмоторный всепогодный тактический истребитель, разработанный McDonnell Douglas (ныне Boeing) для завоевания и поддержания господства в воздухе в воздушных боях.После рассмотрения предложений ВВС США. | General Dynamics F-16 Fighting Falcon – это одномоторный многоцелевой истребитель, первоначально разработанный General Dynamics для ВВС США (USAF). |
Развитие
F-15 был разработан в ответ на стремление ВВС создать доминирующий истребитель превосходства в воздухе, способный наносить удары по земле, частично для противодействия предполагаемой угрозе со стороны советского самолета Mig Foxbat, разрабатываемого в конце 1960-х годов.Требования к высокой скорости / дальности полета привели Макдоннелла Дугласа к двухмоторной конструкции F-15. Сейчас существует почти 10 версий F-15 с такими вариантами, как пониженная радиолокационная заметность, двухместные тренажеры и улучшенные всепогодные модели радаров. Ожидается, что F-15 будет находиться на вооружении ВВС США как минимум до 2025 года.
После войны во Вьетнаме американские военные решили, что им нужен высокоманевренный и недорогой истребитель в дополнение к более мощному и дорогостоящему F-15.F-16 был разработан General Dynamics с целью иметь возможность маневрировать с минимальными потерями энергии и иметь высокое соотношение тяги к весу. Наличие одного двигателя обеспечивало легкость и маневренность самолета. Существует также около 10 вариантов F-16, которые, как ожидается, будут находиться на вооружении ВВС США как минимум до 2025 года.
Посмотрите документальный фильм о F-16 Fighting Falcon:
Характеристики
F-15 отличается своей двухдвигательной конфигурацией, которая дает самолету такую большую тягу, что он может ускоряться, поднимаясь прямо вверх под углом 90 градусов, и может набирать высоту 30 000 футов всего за 60 секунд.Конструкция со сдвоенным хвостом и стреловидным крылом обеспечивает большой угол атаки и хорошую устойчивость на очень высоких скоростях.
F-16 – это односторонний одномоторный самолет, использующий тот же реактивный двигатель Pratt and Whitney P100, что и F-15. Это был первый серийный самолет, спроектированный с пониженной или отрицательной устойчивостью. Большинство самолетов спроектированы с положительной стабильностью, что означает, что они, естественно, хотят вернуться к прямому и горизонтальному полету без участия пилота. Таким образом, ослабление устойчивости означает более эффективное маневрирование с точки зрения потери энергии, поскольку планер не сопротивляется маневрам.Бортовой компьютер помогает поддерживать устойчивость F-16. При полной заправке топливом самолет может выдержать маневры 9G.
F-15 и F-16 летят строем.Сильные стороны
Немногие истребители имеют шанс конкурировать с F-15 по высокой скорости и практическому потолку. Самолет обладает большой дальностью полета, чему способствует возможность установки внешних топливных баллонов, и имеет надежность двух двигателей. Усовершенствованная авионика позволяет ему преследовать врагов с больших расстояний и в плохую погоду.
F-16 очень маневренный и мало теряет энергию при маневрировании, что частично достигается за счет проводной системы управления.Самолет легкий, относительно дешевый и оказался чрезвычайно универсальным. При стоимости около 18 миллионов долларов за единицу он намного дешевле в сборке и эксплуатации, чем F-15.
F-15 на малой высоте, летящий над Северным Уэльсом, и F-16, выполняющий задание в Ираке.Недостатки
Высокие характеристики F-15 обходятся дорого: 30 миллионов долларов за единицу. Во время его разработки критики беспокоились, что самолет был слишком большим и тяжелым, чтобы быть успешным истребителем с собаками, хотя его послужной список доказал обратное.Самолет способен летать быстрее, чем G-рейтинг его планера, что требует наличия системы предупреждения для пилотов, чтобы они не создавали слишком большую нагрузку на самолет.
Поскольку F-16 имеет только один двигатель, он более уязвим для катастрофы двигателя, чем F-15, который может летать с одним двигателем. F-16 также имеет более низкую максимальную скорость, практический потолок и меньшую скорость набора высоты, чем F-15.
F-15 против F-16 Воздушный бой
Интересный воздушный бой между F-15 Eagle и F-16 Falcon:
Список литературы
Поделитесь этим сравнением:
Если вы дочитали до этого места, подписывайтесь на нас:
“F-15 против F-16 Fighting Falcon.” Diffen.com. Diffen LLC, n.d. Web. 19 июня 2021 г. <>
Описание и сравнение микробиоты кожи и слухового прохода неаллергических и аллергических немецких овчарок с использованием секвенирования нового поколения
Реферат
Атопический дерматит – одно из самых распространенных кожных заболеваний у собак. Патогенез сложен и не до конца изучен. Бактерии, колонизирующие кожу, вероятно, играют важную роль в тяжести этого заболевания. Изучение микробиоты кожи собак с использованием традиционных микробиологических методов имеет множество ограничений, которые можно преодолеть с помощью молекулярных процедур.Целью этого исследования было описание бактериальной микробиоты кожи и слуховых проходов здоровых неаллергических и аллергических немецких овчарок (GSD) без острого обострения или сопутствующей кожной инфекции, а также сравнение обоих. Данные о последовательности ампликона бактериального гена 16S рРНК не выявили различий в паттернах бактериального сообщества на разных участках тела (подмышечная впадина, передняя дорсальная межпальцевая кожа, пах и слуховые проходы) у неаллергических собак. Микробиота на разных участках тела неаллергических GSD не показала существенных различий.Только для образцов, полученных из подмышечной впадины, бактериальная микробиота собак с аллергией характеризовалась меньшим видовым разнообразием по сравнению с таковой у неаллергических собак, а состав бактериального сообщества кожи и слуховых проходов собак с аллергией демонстрировал специфические различия в участках тела по сравнению с неаллергические собаки. Актинобактерии были наиболее многочисленным типом, идентифицированным у собак, не страдающих аллергией, а протеобактерии – у собак, страдающих аллергией. Macrococcus spp. были более многочисленны на неаллергической коже, тогда как Sphingomonas spp.были более обильными на аллергической коже. Предварительный поэтапный анализ избыточности метаданных показал, что домохозяйство, из которого произошли собаки, оказало самое сильное влияние на состав микробиома кожи, за которым следовали пол, состояние здоровья хозяина и местонахождение тела.
Образец цитирования: Апостолопулос Н., Глэзер С.П., Багве Р., Янссен С., Майер У., Эверс С. и др. (2021) Описание и сравнение микробиоты кожи и слухового прохода неаллергических и аллергических немецких овчарок с использованием секвенирования следующего поколения.PLoS ONE 16 (5): e0250695. https://doi.org/10.1371/journal.pone.0250695
Редактор: Кристин Мюльдорфер, Институт исследований зоопарков и дикой природы им. Лейбница (IZW), ГЕРМАНИЯ
Поступила: 23 апреля 2020 г .; Одобрена: 13 апреля 2021 г .; Опубликовано: 3 мая 2021 г.
Авторские права: © 2021 Apostolopoulos et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все файлы данных последовательности ампликонов доступны из базы данных SRA с номерами доступа BioSample от SAMN14565128 до SAMN14565223 в BioProject PRJNA624030.
Финансирование: Автор (ы) не получил специального финансирования для этой работы.
Конкурирующие интересы: Один из авторов (UM) работает в коммерческой компании (AniCura Kleintierspezialisten Augsburg GmbH). Эта компания обеспечивала оплату клинической работы автора (УМ).Ее клинические обязанности не зависят от исследования. Более того, у компании не было никакой дополнительной роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи. Конкретная роль этих авторов сформулирована в разделе «Авторские материалы». Эта коммерческая принадлежность не влияет на нашу приверженность политике PLOS ONE в отношении обмена данными и материалами.
Введение
Несколько исследований секвенирования следующего поколения (NGS) за последние несколько лет показали, что кожа собак, как и человеческая, содержит большее разнообразие бактериальных таксонов, чем считалось ранее [1–4].Бактерии играют важную роль как в здоровье, так и в болезнях, а изменения в составе бактериального сообщества кожи связаны со многими кожными заболеваниями как у людей, так и у животных [2,5].
Атопический дерматит собак (САД) – распространенное кожное заболевание у собак, характеризующееся генетически предрасположенным воспалительным, IgE-ассоциированным зудящим аллергическим заболеванием, поражающим определенные участки тела и слуховые проходы [6,7]. ИБС была предложена в качестве животной модели атопического дерматита человека [8,9]. Экологические аллергены являются наиболее частой причиной САД [6].Но пищевые аллергены (кожные побочные реакции на пищу; CAFR) также могут вызывать идентичные клинические признаки или быть фактором обострения КАД, что делает невозможным клиническую дифференциацию [10,11]. Окончательный диагноз должен быть получен путем систематического обследования [12]. Исследования на собаках показали связь между микробиотой кожи и аллергическими кожными заболеваниями. Кожа шести собак с аллергией без признаков пиодермии или дерматита Malassezia показала более низкое разнообразие бактериального сообщества по сравнению с кожей 12 здоровых собак, но состав бактериального сообщества существенно не отличался [13].Недавно проведенное лонгитюдное исследование показало снижение разнообразия и различный состав бактериального сообщества у собак с ХБП и вторичной пиодермией по сравнению со здоровыми собаками [14]. После антимикробной терапии и ремиссии кожных поражений разнообразие было восстановлено, а разница в кластеризации бактериальных сообществ уменьшилась [14]. Ни в одном из исследований не изучали слуховой проход, который часто является участком тела собак, страдающих аллергией. Единственное исследование с использованием NGS для оценки микробиоты бессимптомных слуховых проходов собак с ХБД не показало различий в разнообразии, но существенное различие в составе сообщества бактерий [1].Во всех ранее упомянутых исследованиях участвовали собаки разных пород. Хорошо известно, что САД имеет предрасположенность к породам и что фенотип заболевания различается между породами [15]. Неясно, влияют ли различные фенотипы САД на состав бактериального сообщества между породами. На сегодняшний день ни одно исследование не оценивало микробиоту кожи и слухового прохода только одной породы как при здоровых, так и при аллергических заболеваниях кожи, что минимизировало потенциальную систематическую ошибку из-за влияния фенотипа аллергии на микробиоту.Немецкая овчарка (GSD) была выбрана в нашем исследовании, потому что это порода с высоким риском CAD [15], возможно, из-за измененной экспрессии гена плакофилина 2 и других генов хромосомы 27 [16].
Целью исследования было описание и сравнение бактериальных микробиомов четырех участков тела (подмышечная впадина, передняя дорсальная межпальцевая кожа, пах и слуховой проход) здоровых (неаллергических) и аллергических GSD с использованием гена 16S рРНК на основе ампликона Illumina. секвенсорный подход. Мы предположили, что атопический дерматит и / или CAFR влияют на микробиоту кожи и слухового прохода GSD, что приводит к снижению бактериального разнообразия и значительному изменению состава бактериального сообщества.
Материалы и методы
Объекты исследования
Клиническое исследование проводилось в клинике мелких животных Университета Юстуса Либиха (JLU), Гиссен, Германия. Образцы были собраны в клинике для мелких животных (JLU) во время назначений специально для этого исследования. Все владельцы были проинформированы о процедурах и подписали форму согласия на сбор образцов. Комитет по защите животных Гиссенского университета имени Юстуса Либиха был проинформирован о протоколе исследования и, в частности, обсуждался метод отбора проб.Поскольку мягкое прикосновение ватной палочки к коже не причиняет ни боли, ни вреда, ни ущерба, они заверили нас, что этическое одобрение ответственного органа не требуется. Были изучены две группы GSD. В контрольную группу вошли 12 GSD без каких-либо аллергических состояний в анамнезе или каких-либо клинических поражений кожи / слухового прохода, совместимых с аллергией (в остальной части рукописи они будут называться «неаллергическими») на момент исследования и отбора проб. Чтобы свести к минимуму риск включения собаки с субклинической аллергией, в исследование были включены только собаки старше четырех лет, поскольку аллергические состояния чаще всего развиваются у молодых собак в возрасте от 6 месяцев до 3 лет [17].Чтобы исследовать возможное влияние домашних условий на микробиоту, были включены два неаллергических GSD на одно домашнее хозяйство. За шесть месяцев до отбора проб нельзя было принимать системные антибиотики, какие-либо иммуномодулирующие или противовоспалительные препараты. Купание с шампунем и использование средств для чистки ушей не разрешалось за семь дней до отбора проб. У двенадцати взрослых аллергических GSD был диагностирован CAD, вызванный кожными пищевыми реакциями или аллергенами окружающей среды. Использовались стандартные диагностические и терапевтические методы, включая выполнение по крайней мере пяти критериев Фавро и исключение других зудящих дерматозов (например.грамм. гиперчувствительность слюны блох, саркоптоз) [12,18]. Допускалась комбинация CAD и / или CAFR с гиперчувствительностью слюны блох. Исключались собаки с какой-либо вторичной бактериальной или грибковой инфекцией кожи или уха. Системные и местные антибактериальные или противогрибковые средства не допускались за 30 и 14 дней до отбора образцов соответственно. Системное введение любых иммуномодулирующих или противовоспалительных препаратов, за исключением оклацитиниба, также не разрешалось за два месяца до отбора проб.Местные иммуномодулирующие или противовоспалительные препараты должны были быть отменены за 14 дней до исследования. Мытье шампунем и чистящие средства для ушей не разрешалось за семь дней до отбора проб. Все владельцы заполнили анкету относительно жилья их собаки, животных-партнеров, текущих заболеваний и лечения, пищевых добавок, частоты купания и типа шампуня. Владельцев собак, страдающих аллергией, спрашивали о желудочно-кишечных признаках и о том, как оценить зуд у собак с помощью визуально-аналоговой шкалы зуда (pVAS), как это было ранее подтверждено [19, 20].
Оценка поражений
Все собаки были обследованы клинически и дерматологически, включая отоскопическое обследование с использованием стерильных неопудренных перчаток и автоклавируемого металлического отоскопа. Для атопических собак была проведена валидированная локально-специфическая оценка степени поражения и шкала оценки степени тяжести атопического дерматита у собак (CADESI-4), чтобы оценить любую взаимосвязь микробиоты кожи и тяжести атопического дерматита [ 21]. Цитологию кожи и слухового прохода выполняли, как описано [22,23].
Сбор проб
Перед взятием образцов и между взятием образцов GSD стол для исследования очищали салфетками PCR Clean ™ Wipes (Minerva Biolabs, Берлин, Германия) в соответствии с инструкциями производителя, чтобы избежать перекрестной контаминации ДНК. Чтобы свести к минимуму микробное заражение с пола клиники, собак приносили прямо в комнату для осмотра и помещали на стол для осмотра. После физического обследования были взяты образцы из левой подмышечной впадины (A), левой передней дорсальной межпальцевой области (Int), левой части паха (L) и левого слухового прохода (O).Эти участки тела, по-видимому, наиболее часто поражаются атопией у этой породы [15]. Для выборки микробиомных исследований мы использовали стерилизованные 70% этиленоксидом тампоны для судебно-медицинской экспертизы с транспортной трубкой, материал стержня из полистирола и материал вискозного тампона (Forensic Swab, Nr 80.629, Sarstedt, Nuembrecht, Германия), чтобы гарантировать отсутствие загрязнения ДНК и избежать попадания хлопка или хлопка. древесные митохондрии из тампона или стебля. Мазки натирали 40 раз на желаемом участке, поворачивая одну четверть места тампона (90 °) по 10 раз каждый.Все образцы были получены в двух экземплярах. Впоследствии образец был взят для цитологических целей с помощью стерильного ватного тампона. Образцы были немедленно доставлены при 8 ° C в Институт прикладной микробиологии, JLU Giessen. Образцы для исследований микробиома хранили при -20 ° C до дальнейшей обработки (экстракции ДНК).
Экстракция ДНК и секвенирование ампликона гена 16S рРНК
Тотальную ДНК экстрагировали из мазков без ДНК, хранящихся при -20 ° C, с использованием набора NucleoSpin ® 96 Soil (96-луночная экстракционная система, Macherey Nagel AG, Oesingen, Швейцария), который может эффективно извлекать ДНК из грамотрицательных и грамотрицательных -положительные бактерии, включая споры.Согласно инструкции производителя с использованием вакуумной обработки (вакуумный коллектор NucleoVac 96, Macherey Nagel AG, Oesingen, Швейцария) с небольшими изменениями, экстракция была начата с использованием лизирующего буфера SL1, а затем шаги с 1 по 5 были повторены с лизирующим буфером SL2, таким образом образцы были лизированы дважды. . Тотальную ДНК элюировали 80 мкл воды для ПЦР (1x 30 мкл, 1x 50 мкл) вместо буфера SE. ДНК определяли количественно спектрофотометрически с использованием спектрофотометра NanoDrop (Thermo Scientific), а затем проверяли наличие и возможность амплификации последовательностей гена 16S рРНК для выбранных образцов.
Последовательности гена 16S рРНК бактерий амплифицировали для секвенирования ампликона Illumina (LGC Genomics, Берлин, Германия) с использованием подхода вложенной ПЦР с первой ПЦР с системой праймеров 341F (5′-CCTACGGGAGGCAGCAG-3´) и 1061R (5 ′ -CRRCACGAGCTGACGAC-3´) (V3-V6) [24] (20 циклов) с последующей второй ПЦР с системой праймеров 515F (5′-GTGYCAGCMGCCGCGGTAA-3´) -Y и 926R (5′-CCGYCAATTYMTTTRAGTTT-3´) – jed (V4-V5) [25] (20 циклов), потому что, согласно предыдущим исследованиям, только небольшое количество микробной ДНК было обнаружено на коже человека, проанализированной с помощью ПЦР [26,27].Для каждого образца прямой и обратный праймеры второй ПЦР имели одинаковые последовательности штрих-кода из 10 нуклеотидов. Первый раунд ПЦР проводили в течение 20 циклов, используя следующие параметры: 2 минуты, предварительная денатурация при 96 ° C; Использовались 96 ° C в течение 15 секунд, 50 ° C в течение 30 секунд, 70 ° C в течение 90 секунд и праймеры без встроенных штрих-кодов (341F / 1061R). Для второго раунда использовали 1 мкл продукта ПЦР из первой ПЦР, и условия ПЦР были такими же, как и раньше. В этом случае были добавлены праймеры со штрих-кодом (515F-Y / 926R-jed).Концентрацию ДНК интересующих ампликонов определяли гель-электрофорезом. Около 20 нг ампликонной ДНК каждого образца объединяли до 48 образцов, несущих различные штрих-коды. Пулы ампликонов очищали с помощью гранул AMPure XP одного объема (Agencourt) для удаления димера праймера и других небольших продуктов с ошибочной загрузкой с последующей дополнительной очисткой на колонках MiniElute (Qiagen). Около 100 нг каждой очищенной ДНК пула ампликонов использовали для создания библиотек Illumina с использованием мультиплексной системы Ovation Rapid DR Multiplex System 1–96 (NuGEN).Библиотеки Illumina были объединены и размер выбран препаративным гель-электрофорезом. Секвенирование выполняли на Illumina MiSeq с использованием V3 Chemistry (Illumina). Считывание необработанных последовательностей доступно в Архиве считывания последовательностей (SRA) с регистрационными номерами BioSample от SAMN14565128 до SAMN14565223 в BioProject PRJNA624030.
Анализ данных последовательности ампликона
Для анализа последовательностей использовался конвейер анализа NGS (https://www.arb-silva.de/ngs) базы данных генов рРНК SILVA (SILVAngs 1.3) [28].Для этого в базу данных были загружены наборы данных всех комбинированных считываний последовательностей (обрезанные адаптер и праймер). Все чтения были выровнены с использованием SILVA Incremental Aligner (SINA v1.2.10 для ARB SVN revision 21008) [29], против SILVA SSU rRNA SEED и контроля качества [28]. Считывания короче 50 выровненных нуклеотидов и считывания с более чем 2% неоднозначности или 2% гомополимеров были исключены из дальнейшей обработки. Считывания с низким качеством сопоставления (50 баллов сопоставления, 40 баллов сопоставления по данным SINA), а также предполагаемые загрязнения и артефакты были идентифицированы и исключены из последующего анализа.Следующим шагом процесса была дерепликация и кластеризация с помощью cd-hit-est (версия 3.1.2; http://www.bioinformatics.org/cd-hit) с использованием режима точности , игнорирования выступов и применения критериев идентичности 1,00 и 0,98. соответственно [30]. Для классификации был выполнен локальный поиск нуклеотидов BLAST по неизбыточной версии набора данных SILVA SSU Ref (версия 128; http://www.arb-silva.de) с использованием blastn (версия 2.2.30+; http: // blast.ncbi.nlm.nih.gov/Blast.cgi) со стандартными настройками [31].Уникальные чтения были сгруппированы в операционную таксономическую единицу (OTU) для каждого образца в соответствии с критерием 98% идентичности последовательностей друг другу (попарное расстояние и кластеризация по одиночному сцеплению). Самое длинное чтение в каждом кластере классифицировалось как эталон для каждого OTU и отображалось на все чтения, которые были назначены соответствующему OTU. OTU были отнесены к таксономическим путям (уровень родов). Таким образом, несколько OTU были отнесены к одному таксономическому пути / роду. В результате этого процесса была получена количественная информация (количество индивидуальных считываний всех OTU на таксономический путь), несмотря на ограничения ПЦР, возможные ошибки техники секвенирования и множественные опероны рРНК.Считывания без каких-либо совпадений BLAST или считывания со слабыми совпадениями BLAST, где функция «(% идентичности последовательностей +% покрытия выравнивания) / 2» не превышала значения 93, оставались неклассифицированными и были отнесены к виртуальной таксономической группе «Без относительной» в диаграммах отпечатков пальцев SILVAngs и Krona [32], как сообщалось ранее [33,34].
Анализ альфа- и бета-разнообразия был проведен в PAST3 (https://folk.uio.no/ohammer/past) [35] на уровне родов (филогенетических групп). Альфа-разнообразие измеряли путем расчета различных индексов разнообразия с учетом количества филогенетических групп на образцы и количества прочтений на филогенетическую группу.Были рассчитаны индексы Шеннона (общее альфа-разнообразие) и Chao1 (богатство; количество таксонов, скорректированных на наличие синглтона). Значительные различия между индексами альфа-разнообразия, определенными для неаллергических и аллергических собак, были дополнительно оценены. Для двух групп, отдельных участков тела неаллергических собак по сравнению с аллергическими собаками, непарные t-тесты были выполнены в SigmaPlot 13 (Systat Software Inc.). Сначала был проведен тест на нормальность (Shapiro-Wilk) [36]. Если данные прошли тест на нормальность, двустороннее значение p было получено из анализа t-критерия.Если тест нормальности не удался, применялся тест суммы рангов Манна-Уитни для проверки наличия значимых различий. Для более чем двух групп (все участки тела) значимые различия значений альфа-разнообразия оценивали с помощью теста Краскела-Уоллиса [37]. Бета-разнообразие (сравнение филогенетического состава бактериальных сообществ различных образцов) изучалось на основе данных об относительной численности, проанализированных с помощью неметрической многомерной шкалы (NMDS; [38]) на основе матрицы сходства Брея-Кертиса [39] .Переменные среды, например участки тела, дом (проживание в одном доме) и пол были включены в график NMDS, отображаемый в виде вектора двух графиков. Односторонние ANOSIM (9999 перестановок) [39] и Односторонние PERMANOVA (9999 перестановок) [40] были выполнены для определения значимых различий (оценка последовательных p-значений Бонферрони) между образцами. Анализ SIMPER (процент сходства) [39], также основанный на матрице сходства Брея Кертиса, использовался для определения среднего процентного вклада различных таксонов в несходство между образцами.На основе моделей относительной численности и вклада в различия между сообществами (анализ SIMPER) был дополнительно изучен выбор бактериальных таксонов в отношении значительных различий в относительной численности у неаллергических и аллергических собак. Т-тест проводился, как описано выше. Для коррекции проверки множественных гипотез значения p были разделены на количество выполненных тестов.
Мы количественно оценили размер эффекта нескольких полей метаданных (метка времени сбора, пол, оклацитиниб, состояние здоровья хоста, идентификатор субъекта хоста), объединив их в линейную модель и выполнив анализ прямой ступенчатой избыточности (RDA), как описано ранее [41], с функции rda и ordiR2step в веганском пакете в R 3.6.1 [42].
Путем систематического тестирования полей метаданных на корреляцию с альфа- (с использованием двух метрик «Шеннон», «chao1») или бета-разнообразием (с использованием метрики «Брей-Кертис» [43]) через Краскал-Уоллис [44], Спирман- [45], корреляция Пирсона [46] (для альфа-разнообразия) и PERMANOVA [40] и ANOSIM [39] (для бета-разнообразия), мы обнаружили, что следующие поля были значимыми по крайней мере в одном из тестов: отметка времени сбора , пол, оклацитиниб, состояние здоровья хозяина, идентификатор субъекта хозяина и идентификатор домохозяйства.
Чтобы проиллюстрировать ковариацию между этими полями метаданных, мы исследовали корреляцию между отдельными переменными метаданных. Категориальные поля метаданных (все, кроме метки времени сбора) сравнивались с использованием модифицированной статистики CramérV [47]). Непрерывные (метка времени сбора) и категориальные ковариаты сравнивались с Welchttest [47]. Полученные соотношения корреляции были визуализированы в виде тепловой карты.
Результаты
Объекты исследования
Всего было отобрано 29 GSD, но пять были исключены в соответствии с критериями исключения (таблица 1 в файле S1).Двенадцать неаллергических (6 мужчин, 1 кастрированный мужчина, 4 интактных женщин и 1 стерилизованная женщина) и двенадцать аллергических (2 интактных мужчин, 1 кастрированный мужчина и 9 интактных женщин) были дополнительно проанализированы (таблица 1 в файле S1). Собаки жили в радиусе не более 131 километра от Гиссена (Германия). Возраст неаллергических собак (среднее значение: 7,5 ± 1,9 года) был значительно выше (t-критерий, p <0,01), чем возраст собак, страдающих аллергией (среднее значение: 4,8 ± 2,2 года) (таблица 2 в файле S1). У всех собак с аллергией наблюдались легкие поражения со средним баллом по CADESI-04, равным шести (диапазон 0–11), и средним баллом по шкале PVAS, равным 4 (диапазон 0–8).Две собаки имели оценку PVAS 7/10 и 8/10, соответственно, только с легкими кожными повреждениями. Шесть аллергических GSD получили меченые дозы оклацитиниба (Apoquel, Zoetis Deutschland GmbH, Берлин, Германия).
Анализ филогенетического состава микробиоты GSD
Всего было получено 4 038 850 парных концевых последовательностей со средней длиной последовательности 373 нуклеотида (нуклеотидов). Вкратце, 2579 последовательностей (0,06%) были отклонены из-за того, что они не прошли контроль качества трубопровода SILVAngs.Наконец, 4 036 271 последовательность (2334–246 525 на образец) были дополнительно проанализированы (таблица 3 в файле S1). Всего 241 114 уникальных прочтений (5,97% окончательно проанализированных последовательностей) были отнесены к OTU. Кроме того, 1 129 221 (27,96%) последовательность (количество считываний последовательностей с 98% идентичностью последовательностей друг другу; определяется как «сгруппированные») и 2 665 936 (66,01%) последовательностей (количество считываний последовательностей со 100% идентичностью друг другу; определяется как «реплики») были назначены OTU. Каждая OTU была классифицирована в базе данных SILVA с таксономическими путями с максимальным разрешением на уровне родов (названных «филогенетическими группами» для безымянных родов).Последовательности гена 16S рРНК хлоропласта, митохондрий и архей были обнаружены с относительной численностью 20,13% (190 605 последовательностей), 0,07% (9430 последовательностей), 0,02% (1064 последовательности) от общего количества проанализированных последовательностей, соответственно (таблица 3 в файле S1. ). Последовательности, которые не соответствовали каким-либо известным таксонам (сходство последовательностей <93% со следующим известным таксоном), были классифицированы как «не относящиеся к родственникам» и имели относительную численность 0,3% (5 131 последовательность) (таблица 3 в файле S1). Последовательности, отнесенные к архей , хлоропластам, митохондриям и никаким относительным группам, были исключены из дальнейших анализов.Только последовательности, относящиеся к домену Bacteria (всего 3 830 041; от 2334 до 232 445 на образец), были дополнительно проанализированы и были установлены на 100% (на образец).
Кожный микробиом неаллергенных немецких овчарок
Разнообразие и богатство бактериальных сообществ в образцах кожи неаллергических GSD.
Разнообразие (индекс Шеннона) и богатство (Chao1) филогенетических групп (родов) на четырех участках кожи тела (подмышечная впадина, межпальцевые, пах, слуховой проход) существенно не различались между участками тела (индекс Крускала-Уоллиса для индекса Шеннона, р = 0.6214; для Chao1, p = 0,4784) (Рис. 1 и Рис. 1 в файле S2).
Рис. 1.
Графики индексов альфа-разнообразия Shannon (a) и Chao1 (b) для четырех исследуемых участков тела неаллергических собак. С помощью тестов Краскела-Уоллиса не было получено значительных различий между участками тела.
https://doi.org/10.1371/journal.pone.0250695.g001
Однако большее количество отдельных образцов из межпальцевой области и слухового прохода имело более однородный состав микробиоты, показывая меньшее богатство (значения Chao1 ; Рис.1 в файле S2).
Микробный состав кожи неаллергических GSD.
Паттерны бактериального сообщества, присутствующие на разных участках тела отдельных неаллергических собак, были проверены с помощью анализа NMDS на основе матрицы расстояний, созданной с помощью метрики разнообразия бета-версии Брея Кертиса. Односторонний анализ ANOSIM (p = 0,17) и односторонний анализ PERMANOVA (p = 0,1808) не выявили существенных различий между четырьмя разными участками тела (подмышечная впадина, межпальцевый, пах и слуховой проход) неаллергических GSD.Была задокументирована вариабельность у всех собак и на всех участках тела. Фактор home оказал наиболее сильное влияние на состав бактериального сообщества (наибольшая относительная длина вектора биплоттинга; рис. 2). Существенные различия в составе сообщества были выявлены при группировке по полу (мужчины и женщины; One way ANOSIM и One way PERMANOVA p <0,001; R = 0,1337). Значительные различия в составе бактериального сообщества наблюдались также при группировке по домам (как One way ANOSIM, так и One way PERMANOVA p <0.003; R = 0,1337; Рис. 2 в файле S2; дома, таблица 1 в файле S1).
Рис. 2. Сравнительный анализ состава бактериального сообщества микробиоты кожи всех неаллергических собак.
Сравнительный анализ паттернов относительной численности микробиоты кожи всех участков тела (разных объектов), выполненный с помощью анализа NMDS на основе матрицы сходства Брея-Кертиса. Пол, местонахождение тела и дом были включены в качестве параметров окружающей среды (биплот). Разные цвета представляют отдельных собак.
https://doi.org/10.1371/journal.pone.0250695.g002
Состав (наиболее распространенные таксоны) кожи неаллергических GSD.
Всего было идентифицировано сорок четыре типа из четырех участков выборки (подмышечная впадина, межпальцевый, пах, слуховой проход), взятых у неаллергических собак, из которых 30 типов имели относительную численность менее 1% (Таблица 4 в S1 Файл). Десять наиболее распространенных типов неаллергических собак представляли собой актинобактерии (средняя относительная численность 29,1 ± 14.7%), затем следуют Proteobacteria (27,4 ± 10,1%), Firmicutes (20,3 ± 18,7%) и Bacteroidetes (10,8 ± 5,4%), Cyanobacteria, Acidobacteria (оба 2,0 ± 1,6%), Chloroflexi (1,8 ± 1,6%), Planctomycetes (1,7 ± 1,4%), Deinococcus-Thermus (1,5 ± 1,2%) и Verrucomicrobia (1,1 ± 1,0%) (рис. 3A и 3B). В образцах из межпальцевой области и слухового прохода больше всего были протеобактерии, за ними следовали Actinobacteria, Firmicutes, Bacteroidetes и Cyanobacteria. В образцах паха преобладали актинобактерии, за ними следовали Proteobacteria, Firmicutes, Bacteroidetes и Cyanobacteria.В образцах из подмышечной впадины наиболее многочисленны Firmicutes, за ними следуют актинобактерии, протеобактерии, Bacteroidetes и Cyanobacteria (рис. 3A и таблица 4 в файле S1).
Рис. 3. Филогенетический состав кожи (подмышечная впадина, межпальцевая, паховая) и микробиота слухового прохода неаллергических (n = 12) и аллергических (n = 12) GSD, разрешенных на уровне филума.
(a) Средняя относительная численность бактериальных типов, выявленных в подмышечной впадине, межпальцевой области, паху и слуховом проходе неаллергических и аллергических собак.(b) Изменение относительной численности (по всем участкам тела) 10 наиболее распространенных типов среди неаллергических и аллергических собак. Звездочки представляют статистическую значимость: * p <0,05; ** p <0,01; *** р <0,001.
https://doi.org/10.1371/journal.pone.0250695.g003
Модели относительной численности родов приведены в таблицах 5a и 5b в файле S1. Анализ совокупной средней относительной численности показал, что роды Macrococcus и Staphylococcus были наиболее многочисленными (> 4%) на коже неаллергических собак (рис. 4 и таблицы 5b, 6a и 6b в файле S1).Были получены различия в участках тела (рис. 4A и таблицы 5b и 7 в файле S1). Кожа подмышечной впадины была в основном колонизирована Macrococcus (средняя относительная численность 13,8 ± 29,8%), за которыми следовали Brevibacterium , Staphylococcus , Clostridium sensu stricto 7, Nocardioides , Sph. В межпальцевой области преобладали Clostridium sensu stricto 7 (средняя относительная численность 3,5 ± 2,7%), за которыми следовали Nocardioides , Pelomonas , Sphingomonas , Vibrionimonas , Psychrobacter6 .В паху в основном колонизировали Staphylococcus (средняя относительная численность 10,8 ± 25,5%), за которыми следовали Corynebacterium 1, Conchiformibius , Nocardioides , Brevibacterium , Macrocromocascus Brevibacterium имела самую высокую среднюю относительную численность (3,7 ± 7,6%) в слуховом проходе, за ней следовали Clostridium sensu stricto 7, Sphingomonas , Psychrobacter , Pelomonas , Flavobacterium Псевдомонас .Впоследствии каждый из четырех таксонов с наивысшей средней относительной численностью на участке тела (подмышечная впадина: Macrococcus; межпальцевый: Clostridium sensu stricto 7; пах: Staphylococcus; ушной канал: Brevibacterium ) оценивался индивидуально на предмет значимости. различное обилие между четырьмя участками тела. Ни один из этих таксонов не имел значительно различающейся относительной численности между четырьмя участками тела (нормальность не удалась; тест Краскела-Уоллиса для подмышечной впадины против межпальцевого против паха против слухового прохода; Macrococcus : p = 0.975; Clostridium sensu stricto 7 : p = 0,085; Staphylococcus : p = 0,288; Brevibacterium : p = 0,406).
Рис. 4. Филогенетический состав кожи (подмышечная впадина, межпальцевая, паховая) и микробиота слухового прохода неаллергических (n = 12) и аллергических (n = 12) GSD, разрешенных на уровне рода.
(a) Среднее относительное количество родов бактерий, идентифицированных в подмышечной впадине, межпальцевой области, паху и слуховом проходе неаллергических и аллергических собак. Изображены все роды с общей относительной численностью> 5%.(b) Относительная численность (на всех участках тела) 10 самых распространенных родов неаллергических и аллергических собак. Звездочки представляют статистическую значимость: * p <0,05; ** p <0,01; *** р <0,001.
https://doi.org/10.1371/journal.pone.0250695.g004
Микробиом кожи немецкой овчарки с аллергией
Разнообразие и богатство бактериальных сообществ в образцах кожи аллергических GSD.
Разнообразие (индекс Шеннона) и богатство (Chao1) филогенетических групп на четырех участках тела (подмышечная впадина, межпальцевые пальцы, пах, слуховой проход) аллергических GSD существенно не различались между участками тела (индекс Крускала-Уоллиса для индекса Шеннона, р = 0.334; для Chao1, p = 0,78) (рис. 3 в файле S2).
Микробный состав кожи аллергических GSD.
Паттерны бактериального сообщества на разных участках тела отдельных собак с аллергией были исследованы с помощью анализа NMDS на основе матрицы сходства, созданной с помощью индекса сходства Брея-Кертиса. Односторонний анализ ANOSIM (p = 0,6) и односторонний анализ PERMANOVA (p = 0,32) не выявили значительных различий между четырьмя разными участками тела (подмышечная впадина, межпальцевый, пах и слуховой проход) аллергических GSD (рис. 4 в S2. Файл).
Чтобы оценить, влияет ли оклацитиниб на состав микробиоты кожи собак с аллергией, мы сравнили составы бактериального сообщества, как описано выше для подгрупп с аллергией на оклацитиниб (n = 6) и без оклацитиниба (n = 6). ) (Рис. 5 в файле S2). Не было получено значительной разницы, предполагая, что оклацитиниб не влияет на состав микробиоты кожи и слухового прохода (односторонний ANOSIM, p-значения: подмышечная впадина: p = 0,2777; межпальцевая: p = 0.2949; пах: p = 0,1494; слуховой проход: p = 0,9022) среди собак с аллергией, учитывая небольшую численность популяции подгруппы. Исходя из этого, мы предполагаем, что оклацитиниб не должен влиять на сравнение аллергических и неаллергических собак.
Состав (наиболее распространенные таксоны) кожи аллергических GSD.
Протеобактерии (средняя относительная численность 35,5 ± 10,9%) и актинобактерии (22,4 ± 10,2%) были наиболее многочисленными типами аллергических GSD и доминировали на каждом из четырех участков выборки (рис. 3A и 3B и таблица 8 в файле S1) .За ними следуют Firmicutes (17,0 ± 14,6%), Bacteroidetes (14,1 ± 5,2%), Cyanobacteria (2,5 ± 3,0%), Planctomycetes (1,8 ± 1,2%), Chloroflexi (1,7 ± 1,5%), Acidobacteria (1,7 ± 1,6%). ), Verrucomicrobia (0,7 ± 0,9%) и Deinococcus-Thermus (0,6 ± 0,7%).
Обзор моделей относительной численности родов, присутствующих в микробиоте на различных участках тела собак, страдающих аллергией, приведен в таблицах 5a и 5b в файле S1. Агрегированные данные об относительной численности показали, что Sphingomonas , Staphylococcus , Clostridium sensu stricto 7 и Nocardioides доминировали в микробиоте кожи аллергических GSD (каждый род> 4% совокупной относительной численности) (рис. 6a и 6b в файле S1).Анализ средней относительной численности на уровне рода выявил некоторые специфические различия участков тела (рис. 4A и таблицы 5b и 7 в файле S1). Clostridium sensu stricto 7 (средняя относительная численность 6,2 ± 7,8%) была самой многочисленной филогенетической группой на коже подмышечной впадины, за ней следовали Staphylococcus , Vibrionimonas , Sphingomonas , Pelomonasides Pelomonas . В межпальцевой коже преобладали Nocardioides (средняя относительная численность 5.3 ± 4,4%), затем следуют Sphingomonas , Pelomonas , Clostridium sensu stricto 7 , Vibrionimonas , Bradyrhizobium , Staphylococcus и Hymenobac. В коже паха преобладали Sphingomonas (средняя относительная численность 4,9 ± 2,4%), за которыми следовали Clostridium sensu stricto 7 , Nocardioides , Porphyromonas , Conchiformibius , Pelomonasim .Наконец, в слуховом проходе у аллергических GSD преобладали Staphylococcus (средняя относительная численность 9,4 ± 25,9%), за которыми следовали Sphingomonas , Clostridium sensu stricto 7, Nocardioides , Vibrionomonasimonas Propionibacterium и Paracoccus .
Сравнительный анализ микробиома кожи неаллергических и аллергических немецких овчарок
Различия в разнообразии и богатстве микробиоты кожи неаллергических и аллергических GSD.
При сравнении разнообразия между неаллергическими и аллергическими GSD не было получено значительных различий для индекса Шеннона для каждого участка тела (таблица 9 в файле S1 и рис. 5). Значительно меньшее богатство бактериального сообщества (индекс Chao1) было определено только на коже подмышечной впадины аллергических GSD (A; индекс Chao1; p = 0,032; таблица 9 в файле S1 и рис. 5).
Рис. 5.
Коробчатые диаграммы индексов альфа-разнообразия Шеннона (общее разнообразие) (a) и Chao1 (b) микробиоты кожи на каждом участке тела (подмышечная впадина, межпальцевая область, пах, слуховой проход) неаллергических и аллергических Немецкие овчарки, на основе секвенирования ампликона гена 16S рРНК Illumina в микробных сообществах.Звездочки представляют статистическую значимость: * p <0,05.
https://doi.org/10.1371/journal.pone.0250695.g005
Различия в составе бактериальных сообществ неаллергических и аллергических GSD.
Изменения в составе бактериального сообщества на коже собак с аллергией по сравнению с неаллергическими собаками были визуализированы на графиках NMDS (рис. 6). Достоверные различия были получены для подмышечной впадины (A; односторонний ANOSIM p = 0,048, R = 0.1001; Односторонний PERMANOVA p = 0,0256), пах (L; Односторонний ANOSIM p = 0,036, R = 0,00386; Односторонний PERMANOVA p = 0,0264) и слуховой проход (O; Односторонний ANOSIM p = 0,0012, R = 0,1842; Односторонний PERMANOVA p = 0,0025), но не для межпальцевой области (Int; односторонний ANOSIM p = 0,1548; односторонний PERMANOVA p = 0,1171). Включение статуса здоровья (неаллергический против аллергического) подтвердило, что аллергия коррелировала со значительными различиями в кластеризации структур сообщества между выборками (рис. 6).Образец a18O (слуховой канал собаки с аллергией a18) был исключен из графика NMDS, потому что сообщество было слишком разным, чтобы более четко визуализировать незначительные различия между другими образцами (рис. 6 в файле S2). Сильное различие профиля сообщества было связано с высокой относительной численностью рода Staphylococcus (91,3%) в этом образце кожи по сравнению с 0,02–6,4% в других образцах кожи (Таблица 5 в файле S1).
Рис. 6. Сравнительный анализ состава микробиоты кожи с моделями относительной численности на каждом участке тела неаллергических (n = 12; черный цвет) и аллергических (N = 12; красный цвет) собак, выполненный с помощью анализа NMDS на основе Матрица подобия Брея-Кертиса.
Состояние здоровья было включено как параметр окружающей среды; (A-подмышечная впадина, межпальцевая область, L-пах, ушной канал).
https://doi.org/10.1371/journal.pone.0250695.g006
Чтобы определить, какие таксоны (филогенетические группы / роды) внесли основной вклад в различия в сообществах между неаллергической и аллергической GSD в целом и для для каждого участка тела был проведен анализ SIMPER (таблицы 6a и 10 в файле S1). Staphylococcus и Macrococcus имели 4.89% и 4,8% – самый высокий вклад в общие различия между колонизацией неаллергических и аллергических собак, если объединить отн. были сопоставлены паттерны численности бактериальных родов (анализ SIMPER; таблица 6 в файле S1). Роды с наибольшим вкладом в различия в сообществах среди неаллергических и аллергических собак (> 2%) по отдельным участкам тела перечислены в Таблице 10 в файле S1. Род Macrococcus (вклад 9,0%) внес основной вклад в различия в образцах из подмышечной впадины (A), за ним следует Staphylococcus (4.7% взнос). Для образцов из паха (L) род Staphylococcus внес наибольший вклад (вклад 7,0%). Для слухового прохода (O) наибольший вклад в различия (вклад 2,5%) внес Brevibacterium , при этом Staphylococcus из образца a18O исключены как преобладающие среди всех остальных, как упоминалось выше, влияющих на сравнение.
Состав (наиболее распространенные таксоны) кожи неаллергических и аллергических GSD.
Порядок 10 наиболее распространенных типов аллергических GSD отличался от неаллергических GSD (таблицы 4 и 8 в файле S1; рис. 3).Между двумя группами Proteobacteria (p <0,001) и Bacteroidetes (p = 0,003) имели значительно более высокую среднюю относительную численность на коже собак с аллергией, тогда как Actinobacteria (p = 0,012), Deinococcus-Thermus (p <0,001) и Verrucomicrobia. ( p = 0,016) встречались в значительно более низкой относительной численности у собак с аллергией. Фирмикуты не показали значительных различий между аллергическими и неаллергическими GSD (рис. 3B).
Оценка средней относительной численности филогенетических групп для значимых различий между неаллергическими и аллергическими GSD показала, что Sphingomonas были значительно более распространены среди аллергических GSD (p <0.001; средний отн. обилие неаллергических собак 2,5 ± 1,4% против 4,9 ± 2,3% собак с аллергией), а также Nocardioides (р = 0,034; среднее относительное содержание 2,5 ± 1,8% для неаллергических собак против 4,3 ± 3,4% для аллергические собаки) (рис. 4В). Для Staphylococcus (p = 0,8, таблица 11a в файле S1) и Clostridium sensu stricto 7 (p = 0,062) между неаллергическими и аллергическими GSD не наблюдалось значительной разницы. Интересно, что у собак с аллергией средняя относительная численность Macrococcus была значительно ниже (p <0.001; средний отн. обилие 4,8 ± 15,5% для собак, не страдающих аллергией, против 0,1 ± 0,4% для собак, страдающих аллергией; Таблица 11b в файле S1) и Brevibacterium (p = 0,016; среднее относительное содержание 2,7 ± 6,3% для собак без аллергии по сравнению с 0,1 ± 0,3% для собак с аллергией), чем неаллергические GSD.
Более подробное сравнение неаллергических и аллергических GSD наиболее распространенных таксонов в зависимости от участка тела выявило важные результаты (рис. 4). Clostridium sensu stricto 7 встречался в значительно большей относительной численности в подмышечной впадине собак с аллергией (p = 0.026; средняя относительная численность 2,7 ± 5,0% для собак, не страдающих аллергией, по сравнению с 6,2 ± 7,8% для собак, страдающих аллергией). Nocardioides межпальцевой кожи не показали значимых различий (p = 0,138). Sphingomonas было значительно больше в паху собак с аллергией (p = 0,002; средняя относительная численность 2,0 ± 1,5% для собак без аллергии по сравнению с 4,9 ± 2,4% для собак с аллергией). Кроме того, Sphingomonas были значительно более многочисленными в неаллергических по сравнению с аллергическими GSD для нескольких участков (подмышечная впадина: p = 0.013; межпальцевые: p = 0,014; слуховой проход: p = 0,017, таблица 7 в файле S1). Staphylococcus не обнаружил каких-либо существенных различий ни в одном месте между аллергическими и неаллергическими GSD (таблица 11a в файле S1). Напротив, значительно меньшая относительная численность Macrococcus была получена из образцов межпальцевой кожи, паха и слухового прохода собак с аллергией, но не было значительных различий для подмышечной впадины (таблица 11b в файле S1). Кроме того, Brevibacterium также оценивали на предмет значительных различий в образцах слухового прохода, поскольку он внес основной вклад в разницу в составе бактериального сообщества между двумя группами в анализе SIMPER (таблица 10 в файле S1) с аллергической группой. имея значительно более низкую среднюю относительную численность (p = 0.041; среднее относительное содержание микробиоты ушного канала (таблица 7 в файле S1) составляет 3,6 ± 7,7% для неаллергических собак по сравнению с 0,1 ± 0,4% для аллергических собак).
Оценка влияния метаданных на анализ альфа- и бета-разнообразия микробиоты кожи
Наконец, влияние метаданных (отметка времени сбора, пол, оклацитиниб, состояние здоровья хозяина, идентификатор субъекта-хозяина) на данные об альфа- и бета-разнообразии для всех изученных собак и участков тела было оценено с помощью анализа RDA. Мы обнаружили, что следующие поля были значимыми, по крайней мере, в одном из тестов: отметка времени сбора, пол, оклацитиниб, состояние здоровья хозяина, идентификатор объекта хозяина и идентификатор домохозяйства.На значения индекса альфа-разнообразия (Шеннон, Чао1) существенно повлияло только домохозяйство (рис. 7 в файле S2), и если этот фактор был исключен, только пол собак показал значительный вклад в альфа-разнообразие (рис. 7 в файле S2). Для бета-разнообразия было получено большее влияние различных факторов. Здесь снова домохозяйство оказало преобладающее влияние на различия в микробиоте кожи в сообществе (рис. 7A). Менее значимое влияние наблюдалось для факторов: место расположения тела хозяина и отметка времени сбора.Если фактор домохозяйства был исключен из анализа RDA для бета-разнообразия, пол, за которым следовала отметка времени сбора, состояние здоровья собаки и место на теле хозяина оказали в убывающем порядке влияние на различия в составе бактериального сообщества (рис. 7B). Взаимосвязь различных метаданных проиллюстрирована на рис. 7C. Домохозяйство в сочетании с идентификатором субъекта-хозяина (отдельные собаки), состоянием здоровья хозяина и лечением оклацитинибом оказали сильнейшую взаимосвязь и влияние на изменения в сообществе микробиоты кожи.
Рис. 7.
Анализ прямой ступенчатой избыточности (RDA), показывающий влияние метаданных на анализ бета-разнообразия, включая все изученные собаки и участки кожи тела (a; b, за исключением фактора домохозяйство) и тепловую карту, показывающую взаимосвязь метаданных (c) .
https://doi.org/10.1371/journal.pone.0250695.g007
Обсуждение
В этом исследовании NGS мы описали бактериальное сообщество кожи и слухового прохода неаллергических GSD и сравнили его с микробиотой кожи и слухового прохода аллергических GSD.В нескольких исследованиях NGS также изучалась кожа собаки или слуховой проход в состоянии здоровья и болезни. В этих исследованиях использовались различные методы, от сбора и хранения образцов до выделения и анализа ДНК, и, как было показано ранее, методология влияет на результаты [48–50]. Поэтому прямое параллельное сравнение исследований затруднено [48]. Тем не менее, любой из этих результатов полезен для дальнейшей характеристики микробиоты кожи собак в состоянии здоровья и болезни. Наш подход к ампликону гена 16S рРНК (вложенная ПЦР; V3-V6 и V4-V5) показал высокую индивидуальную вариабельность и вариабельность участков тела у разных испытуемых без значительных различий между участками тела, в отличие от первого ветеринарного исследования на основе NGS. медицина [13].В нашем исследовании кожа тела (подмышечная впадина, межпальцевая область, пах) показала значительно большее разнообразие видов, чем ушной канал у неаллергических собак. Возможные объяснения могут включать особую конструкцию слухового прохода (камерную) и гистологию со сравнительно тонким эпидермисом и дермой, содержащими отдельные волосяные фолликулы, сальные и церуминальные железы, обеспечивающие богатую липидами серную пыль, определяющую микросреду с относительно высокой влажностью. [51,52]. Бактерицидная активность серы может объяснить меньшее видовое богатство по сравнению с другими участками тела.
Мы определили актинобактерии как наиболее доминирующий тип у неаллергических собак, как и в другом исследовании [53]. Этот вывод также согласуется с бактериальным составом кожи человека [3]. Напротив, первое ветеринарное исследование NGS, описывающее бактериальный состав кожи здоровых собак, показало преимущественно протеобактерии [13]. Как упоминалось выше, различные методологии могут объяснить такие различия, как показали исследования [48,50,51,54]. Таким образом, это явление характерно не только для собак.Кроме того, исследования показали, что факторы окружающей среды могут влиять на микробиом кожи человека, например, контакт с почвой или растительным материалом [53,55–57]. Следовательно, можно предположить, что различия в результатах могут быть объяснены разными привычками или средой обитания собак. Кроме того, временные изменения микробиома также могут влиять на результаты и приводить к таким разногласиям между исследованиями. На сегодняшний день временная стабильность кожного микробиома животных остается в значительной степени неизвестной. У крыс здоровая кожа показала временную нестабильность (11 дней) [58], тогда как здоровый слуховой проход собаки оставался стабильным в течение 28 дней.[1]. В соответствии с другими исследованиями, проживание в одном доме и пол повлияли на структуру бактериального сообщества в выборках здоровых собак [53,59]. Интересно, что в нашем исследовании Proteobacteria преобладали в ушном канале и межпальцевой области, как и в другом исследовании межпальцевой области и ушной раковины [13]. Непонятно, почему эти две области показали такую разницу по сравнению с паховой и подмышечной областями GSD. Одним из возможных объяснений межпальцевой области может быть прямой и многократный контакт с почвой, где показано, что в ее бактериальном составе преобладают протеобактерии [60].В настоящее время есть только два других исследования NGS, описывающих слуховой проход собаки. В первом исследовании Proteobacteria были наиболее многочисленными типами, за ними следовали Actinobacteria, Firmicutes, Bacteroidetes и Fusobacteria с наиболее распространенным родом Escherichia [1]. Второе исследование также показало аналогичные наиболее распространенные типы, но в другом порядке: Firmicutes были наиболее многочисленными, за ними следовали Proteobacteria, Bacteroidetes и Actinobacteria с наиболее многочисленным родом Romboutsia [61].Наши результаты согласуются с первым исследованием [1], поскольку у нас был тот же порядок наиболее распространенных типов, за исключением того, что мы задокументировали цианобактерии вместо фузобактерий. На уровне рода мы определили Brevibacterium как наиболее распространенный таксон. Самым многочисленным видом, обнаруженным на неаллергической коже, был род Macrococcus с наибольшей численностью в подмышечной впадине. Вторым по распространенности был род Staphylococcus с наибольшей численностью в паху, за которым следуют подмышечная впадина, межпальцевая область и слуховой проход. Clostridium sensu stricto 7 – самый распространенный род межпальцевой области. Напротив, одно предыдущее исследование показало, что здоровая кожа собаки (подмышечная впадина, ушная раковина и пах) и во рту были преимущественно колонизированы Porphyromonas , Staphylococcus , Streptococcus и Propionibacterium , причем Porphyromonas в значительной степени колонизировали аксиллярные колонии. [14]. В другом исследовании анализ здоровой кожи собак (спинной нос, слизистая оболочка носа, спайка губ, конъюнктива, периокулярная кожа, ушной канал, вогнутая ушная раковина, дорсальная поясничная область, подмышечная впадина, пах, межпальцевая кожа и перианальная область) выявили Ralstonia как наиболее многочисленный род в большинстве выборок [13].Недавнее исследовательское исследование с участием шести здоровых собак показало, что на коже (паховой, подмышечной, периокулярной и туловищной) наиболее распространено Pseudomonas , за которыми следуют Kocuria , Porphyromonas и Corynebacterium [62]. В то время как другой идентифицировал Propionibacterium acnes , Corynebacterium и Porphyromonas как наиболее распространенных в коже (дорсальная часть шеи, подмышечная впадина и живот) 40 здоровых собак [53]. Эти результаты противоречат нашему исследованию. Macrococcus – грамположительная коккоидная бактерия, ранее относившаяся к роду Staphylococcus , но с 1998 г. отнесенная к собственному роду [64]. Он состоит из восьми видов, тесно связанных с видами рода Staphylococcus [63]. Текущая информация о распространении Macrococcus ограничена. Этот род описан в первую очередь как часть микробиоты млекопитающих, а также в молоке и мясе согласно текущему обзору [63].Несмотря на то, что она считается непатогенной бактерией, имеется несколько сообщений об инфекциях, связанных с Macrococcus caseolyticus и M . canis [64–67]. Недавнее исследование показало, что из 162 собак 13 несли M . canis и шесть M . caseolyticus преимущественно в кожных (подмышечных и паховых) неинфекционных участках. Шесть M . canis и один M . caseolyticus. штаммов были выделены от животных с ринитом, наружным отитом, дерматитом и маститом [64].Поскольку была колонизирована как здоровая, так и инфицированная кожа, был сделан вывод, что Macrococcus является важной условно-патогенной бактерией кожи собаки [64]. Поскольку Macrococcus способствовал, главным образом, различию в составе бактериального сообщества подмышечной впадины между неаллергическими и аллергическими GSD, и был родом с самым высоким относительным содержанием, обнаруженным на коже неаллергических собак, можно предположить потенциальную защитную роль, тоже. Таким образом, необходимы дальнейшие исследования распространения и роли бактерий этого рода у здоровых и больных собак.
Важным открытием нашего исследования было то, что местно-специфический бактериальный состав значительно различается при аллергическом кожном заболевании. Анализ SIMPER выявил таксоны, которые вносят основной вклад в эти изменения, хотя средняя относительная численность некоторых из этих таксонов (например, Staphylococcus ) существенно не различалась между неаллергическими и аллергическими GSD. Возможное объяснение этого заключается в том, что анализ SIMPER выявил роды, которые имели наибольший вклад в различия между группами, но не обязательно существенно различались.Кроме того, подмышечные впадины собак с аллергией показали значительно уменьшенное разнообразие (видовое богатство), указывающее на дисбактериоз (из-за более высокой относительной численности Clostridium sensu stricto 7 и Sphingomonas ), хотя у атопических собак не было клинических обострений и не было пиодермии во время отбора проб. Это согласуется с другим исследованием атопических собак с пиодермией, которое продемонстрировало различные бактериальные сообщества и снижение разнообразия у собак с аллергией [14].Первое исследование NGS на основе ампликона гена 16S рРНК с участием собак с аллергией без обострения, как и в нашей исследуемой популяции, не показало значительных различий в составе сообщества микробиоты кожи по сравнению со здоровыми собаками в отличие от наших результатов, но показало, что аллергическая кожа имела более низкую разнообразие (богатство), аналогично нашим выводам [13]. Ушной канал собак с аллергией без признаков отита также имел бактериальный состав, значительно отличающийся от ушного канала здоровых собак, и без существенной разницы в разнообразии между группами, что аналогично нашим результатам [1].Тенденция к развитию дисбактериоза в сочетании с аллергией была продемонстрирована при значительном увеличении численности Staphylococcus и Ralstonia в атопических слуховых проходах [1]. В целом, мы смогли показать тенденцию к дисбактериозу для всех участков, оцененных в целом, со значительным сокращением Macrococcus и Brevibacterium и увеличением типа Proteobacteria и родов Sphingomonas и Nocardioides .
Шесть из 12 собак с аллергией получали оклацитиниб, который является ингибитором киназы Януса с противозудными и противовоспалительными свойствами [68].До сих пор не проводилось исследований, посвященных влиянию оклацитиниба на микробиоту кожи. Одно исследование документально подтвердило, что лечение циклоспорином или кортикостероидами не влияло на микробиоту кожи у собак, при оценке этих собак до, во время и после лечения [69]. Это согласуется с нашими выводами: бактериальный состав собак с аллергией, получавших и не принимавших оклацитиниб, не показал каких-либо значительных различий. Таким образом, мы предполагаем, что оклацитиниб может не влиять на общее сравнение неаллергических и аллергических собак, но будущие исследования с более широкой популяцией должны подтвердить наши выводы.
Одним из ограничений нашего исследования был небольшой размер выборки. Однако размер нашей выборки больше, чем в предыдущих исследованиях [1,13,62,69]. Кроме того, наше исследование дополняет растущий массив исследований, которые помогают нам лучше понять микробиоту, населяющую кожу и слуховой проход собак, и могут использоваться для разработки более крупных подтверждающих исследований. Исследование представляло собой перекрестный анализ, поэтому остается неясным, являются ли полученные данные причиной или результатом аллергии. Что касается описания микробиоты кожи, только продольное исследование может прояснить, является ли состав микробиоты стабильным или временным.Необходимы дальнейшие исследования, включая также неаллергических собак других пород, чтобы оценить, является ли микробиота кожи неаллергических GSD конкретной породой. Поскольку предыдущие исследования микробиоты кожи человека выявили небольшое количество микробной ДНК при использовании стандартной ПЦР, в нашем исследовании была выбрана вложенная ПЦР [26,27]. Смещение в альфа-разнообразии и структуре сообщества (бета-разнообразие = было задокументировано из-за вложенной ПЦР в образцах стула, но не в вагинальных мазках, и может рассматриваться как возможный фактор ограничения нашего исследования [70].Однако количество микробиоты и ДНК, полученное из кожи и в ушных каналах собак с помощью взятия мазков, больше похоже на результат, полученный из вагинальных мазков, чем из образцов стула, которые, как правило, содержат более высокую нагрузку бактерий / ДНК, которые вызывают вложенный подход к этим образцам не нужен. В настоящее время не существует стандартного протокола для методологии, и, как следствие, в разных исследованиях используются разные праймеры [48]. Поскольку существует очень ограниченное систематическое сравнение праймеров и не существует «идеального» праймера [71], чаще всего праймеры выбираются на основе рекомендаций и экспериментального опыта лаборатории [54].Наша вторая система праймеров нацелена на гипервариабельную область V4-V5, которая в настоящее время используется для микробного профилирования на основе секвенирования гена 16S рРНК [25,54,72]. Наш выбор был основан на исследованиях, которые предлагают точную оценку множества таксонов и эффективное филогенетическое разрешение с этой гипервариабельной областью [25,54]. Эти преимущества были недавно вновь продемонстрированы [73]. С другой стороны, как было показано в том же исследовании, нацеливание на этот регион (или нацеливание на другие регионы) может повлиять на результаты, потому что разные праймеры не всегда обнаруживают ожидаемые специфические бактериальные сообщества, что приводит к смещению при количественной оценке. этих сообществ оценивается [73].Но система праймеров, усиливающая V4-V5, была одной из систем праймеров с менее предвзятым отношением, и, вероятно, эти эффекты могут быть незначительными [73]. Также было показано, что анализ состава бактериального сообщества (исследования бета-разнообразия) был устойчивым для использования различных праймеров [71,73]. Чтобы свести к минимуму возможность включения собаки с субклинической ХБП в неаллергическую группу, возраст здоровых собак должен быть более четырех лет. Следовательно, возраст собак двух групп существенно отличался.Это может быть ограничением, поскольку на людях было показано, что микробиота кожи эволюционирует с возрастом (в первые годы после рождения или с половым созреванием) [74–76]. Кроме того, все собаки жили в одном районе Германии, и окружающая среда влияет на микробиоту кожи [77], что, возможно, приводит к различиям между результатами исследований, проведенных в других странах. В нашем исследовании мы не оценивали связь сельской или городской среды с микробиотой кожи собак. Последним ограничением нашего исследования является то, что секвенирование гена 16S рРНК не может отличить мертвые бактерии от живых [2].Более крупные исследования с участием других пород, нескольких стран, а также собак с обострениями аллергии и без них могут пролить свет на микробиоту кожи собак в состоянии здоровья и болезней.
Заключение
Это исследование описывает и сравнивает микробиоту кожи и слухового прохода неаллергических и аллергических GSD, показывая, что аллергические собаки имеют значительно различающийся состав бактериального сообщества и значительно более низкое видовое богатство (альфа-разнообразие). Микробиота кожи и слухового прохода у неаллергических GSD сильно варьируется, и слуховой канал имеет значительно меньшее видовое богатство, чем кожа тела.
Благодарности
Мы благодарим Катю Гребинг и Анну Баум (прикладная микробиология, JLU Giessen) за их отличную техническую помощь.
Ссылки
- 1. Ngo J, Taminiau B, Fall PA, Daube G, Fontaine J. Микробиота слухового прохода – сравнение здоровых собак и собак с атопией без клинических признаков наружного отита. Vet Dermatol. 2018; 29: 425 – e140. pmid: 30084115.
- 2. Родригес Хоффманн А. Кожная экосистема.Роль микробиома кожи в здоровье и его связь с воспалительными состояниями кожи у людей и животных. Vet Dermatol. 2017; 28: 60 – e15. pmid: 28133874.
- 3. Костелло Е.К., Лаубер К.Л., Хамади М., Фирер Н., Гордон Дж. И., Найт Р. Вариации бактериального сообщества в средах обитания человеческого тела в пространстве и времени. Наука. 2009; 326: 1694–7. pmid: 19892944.
- 4. Грайс Э.А., Сегре Я. Микробиом кожи. Nat Rev Microbiol. 2011; 9: 244–53. pmid: 21407241.
- 5. Каркман А., Лехтимяки Дж., Руоколайнен Л. Экология микробиоты человека: динамика и разнообразие в состоянии здоровья и болезней. Ann N Y Acad Sci. 2017; 1399: 78–92. pmid: 28319653.
- 6. Холливелл Р. Пересмотренная номенклатура ветеринарной аллергии. Вет Иммунол и Иммунопатол. 2006; 114: 207–8. pmid: 17005257.
- 7. Хиллиер А., Гриффин К.Э. Рабочая группа ACVD по атопическому дерматиту собак (I): частота и распространенность. Вет Иммунол и Иммунопатол.2001; 81: 147–51. pmid: 11553375
- 8. Freudenberg JM, Olivry T, Mayhew DN, Rubenstein DS, Rajpal DK. Сравнение транскриптомов кожи подтверждает, что атопический дерматит собак является естественным гомологом болезни человека. J Invest Dermatol. 2018. pmid: 30393079.
- 9. Марселла Р., Джироломони Г. Собачьи модели атопического дерматита: полезный инструмент с неиспользованным потенциалом. J Invest Dermatol. 2009; 129: 2351–7. pmid: 19516261.
- 10. Джексон HA, Мерфи KM, Tater KC, Olivry T, Hummel JB, Itensen J, et al.Модель гиперчувствительности к аллергенам (диетическая или экологическая) у собак с несезонным атопическим дерматитом не может быть дифференцирована на основе исторической или клинической информации: проспективная оценка 2003–2004 гг. Выдержки из совместного заседания Американского колледжа ветеринарной дерматологии и Американской академии ветеринарной дерматологии 6–10 апреля 2005 г., Сарасота, Флорида; 16; 2005. С. 192–211.
- 11.
Olivry T, Deboer DJ, Prélaud P, Bensignor E. Пища для размышлений: размышление о взаимосвязи между атопическим дерматитом собак и кожными побочными реакциями на пищу.Vet Dermatol. 2007; 18: 390–1. pmid: 179
. - 12. Хенсель П., Санторо Д., Фаврот С., Хилл П., Гриффин С. Атопический дерматит собак: подробные рекомендации по диагностике и идентификации аллергенов. BMC Vet Res. 2015; 11. pmid: 26260508.
- 13. Hoffmann AR, Patterson AP, Diesel A, Lawhon SD, Ly HJ, Stephenson CE и др. Микробиом кожи здоровых собак и собак с аллергией. PLOS ONE. 2014; 9: e83197. pmid: 24421875
- 14. Брэдли К.В., Моррис Д.О., Ранкин С.К., Каин С.Л., Мисик А.М., Хаузер Т. и др.Продольная оценка микробиома кожи и ассоциации с микросредой и лечением атопического дерматита собак. J Invest Dermatol. 2016; 136: 1182–90. pmid: 26854488.
- 15. Вильхем С., Ковалик М., Фаврот С. Связанные с породой фенотипы при атопическом дерматите собак. Vet Dermatol. 2011; 22: 143–9. pmid: 20887404.
- 16. Tengvall K, Kierczak M, Bergvall K, Olsson M, Frankowiack M, Farias FHG и др. Полногеномный анализ немецких овчарок выявляет ассоциацию локуса CFA 27 с атопическим дерматитом.PLOS Genetics. 2013; 9: e1003475. pmid: 23671420
- 17. Гриффин С., ДеБоер Д. Целевая группа ACVD по атопическому дерматиту собак (XIV): клинические проявления атопического дерматита собак. Ветеринарная иммунология и иммунопатология. 2001; 81: 255–69. pmid: 11553388
- 18. Favrot C, Steffan J, Seewald W, Picco F. Проспективное исследование клинических особенностей хронического атопического дерматита собак и его диагностики. Vet Dermatol. 2010; 21: 23–31. pmid: 20187911.
- 19.Hill PB, Lau P, Rybnicek J. Разработка оцененной владельцем шкалы для измерения тяжести зуда у собак. Vet Dermatol. 2007; 18: 301–8. pmid: 17845617.
- 20. Рыбницек Дж., Лау-Гиллард П. Дж., Харви Р., Хилл ПБ. Дальнейшая валидация шкалы тяжести зуда для использования у собак. Vet Dermatol. 2009; 20: 115–22. pmid: 19171021.
- 21. Olivry T, Saridomichelakis M, Nuttall T, Bensignor E, Griffin CE, Hill PB. Проверка индекса распространенности и тяжести атопического дерматита собак (CADESI) -4, упрощенной шкалы тяжести для оценки кожных поражений атопического дерматита у собак.Vet Dermatol. 2014; 25: 77–85, e25. pmid: 24461108.
- 22. Ginel PJ, Lucena R, Rodriguez JC, Ortega J. Полуколичественная цитологическая оценка нормальных и патологических образцов из наружного слухового прохода собак и кошек. Vet Dermatol. 2002; 13: 151–6. pmid: 12074704
- 23. Будах SC, Мюллер RS. Воспроизводимость полуколичественного метода оценки кожной цитологии. Vet Dermatol. 2012; 23: 426 – e80. pmid: 22809453
- 24. Онг Ш., Куккиллайя В.Ю., Вильм А., Лей С, Ho EXP, Low L и др.Идентификация видов и профилирование сложных микробных сообществ с использованием дробовика Illumina секвенирования последовательностей ампликона 16S рРНК. PLOS ONE. 2013; 8: e60811. Epub 2013/04/08. pmid: 23579286.
- 25. Parada AE, Needham DM, Fuhrman JA. Каждая база имеет значение: оценка праймеров малых субъединиц рРНК для морских микробиомов с помощью фиктивных сообществ, временных рядов и глобальных полевых образцов. Environ Microbiol. 2016; 18: 1403–14. pmid: 26271760.
- 26. Zeeuwen PL, Boekhorst J, Bogaard, Ellen H van den, Koning HDd, Kerkhof, Peter MC van de, Saulnier DM, et al.Динамика микробиома эпидермиса человека после нарушения кожного барьера. Genome Biol. 2012; 13: 1–18. pmid: 23153041
- 27. Ли В, Хан Л., Ю П, Ма Ц, Ву Х, Мур Дж. Э. и др. Молекулярная характеристика микробиоты кожи больных раковой кахексией и здоровых добровольцев. Microb Ecol. 2014; 67: 679–89. pmid: 24402361.
- 28. Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T., Yarza P, et al. Проект базы данных генов рибосомной РНК SILVA: улучшенная обработка данных и веб-инструменты.Nucleic Acids Res. 2013; 41: D590 – D596. pmid: 23193283
- 29. Прюсс Э., Пеплис Дж., Глёкнер Ф. SINA: точное высокопроизводительное выравнивание множественных последовательностей генов рибосомной РНК. Биоинформатика. 2012; 28: 1823–9. pmid: 22556368
- 30. Ли В., Годзик А. Cd-hit: быстрая программа для кластеризации и сравнения больших наборов белковых или нуклеотидных последовательностей. SINA: точное высокопроизводительное выравнивание множественных последовательностей генов рибосомной РНК. Биоинформатика. 2012; 28: 1823–9.pmid: 22556368
- 31. Камачо К., Кулурис Дж., Авагян В., Ма Н., Пападопулос Дж., Билер К. и др. BLAST +: архитектура и приложения. BMC Bioinformatics. 2009; 10: 1–9. pmid: 1
96 - 32. Ондов Б.Д., Бергман Н.Х., Филлиппи А.М. Интерактивная метагеномная визуализация в веб-браузере. BMC Bioinformatics. 2011; 12: 1–10. pmid: 21199577
- 33. Ионеску Д., Зиберт С., Полерецкий Л., Мунвес Ю.Ю., Лотт С., Хойслер С. и др. Микробиологическая и химическая характеристика подводных пресноводных источников Мертвого моря.PLoS ONE. 2012; 7: e38319. pmid: 22679498.
- 34. Клиндворт А., Прюсс Э., Швир Т., Пеплис Дж., Кваст С., Хорн М. и др. Оценка общих праймеров для ПЦР гена 16S рибосомной РНК для классических исследований и исследований разнообразия на основе секвенирования следующего поколения. Nucleic Acids Res. 2013; 41: e1. pmid: 22933715.
- 35. Хаммер Ойвинд, Харпер Дэвид А.Т. и Райан Пол Д. ПРОШЛОЕ: Пакет программного обеспечения палеонтологической статистики для обучения и анализа данных. Palaeontologia Electronica, 4 (1): 9pp.
- 36. Шапиро СС, Вилк МБ. Тест дисперсионного анализа на нормальность (полные образцы). Биометрика. 1965; 52: 591.
- 37. Zar JH. Биостатистический анализ. 3 рд изд. Верхняя Сэдл-Ривер, Нью-Джерси: Прентис-Холл; 1996.
- 38. Taguchi Y-H, Oono Y. Относительные паттерны экспрессии генов с помощью неметрического многомерного масштабного анализа. Биоинформатика. 2005; 21: 730–40. pmid: 15509613.
- 39. CLARKE KR.Непараметрический многомерный анализ изменений в структуре сообщества. Austral Ecol. 1993; 18: 117–43.
- 40. Андерсон MJ. Новый метод непараметрического многомерного дисперсионного анализа. Austral Ecol. 2001; 26: 32–46.
- 41. Thomas RL, Jiang L, Adams JS, Xu ZZ, Shen J, Janssen S и др. Метаболиты витамина D и микробиом кишечника у пожилых мужчин. Nat Commun. 2020; 11: 5997. Epub 2020/11/26. pmid: 33244003.
- 42. веганский: Пакет «Экология сообщества».Пакет R версии 2.4–6. Веганский 2,4–5 (2017).
- 43. Брей Дж. Р., Кертис Дж. Т.. Посвящение горным лесным общинам Южного Висконсина. Экологические монографии. 1957; 27: 325–49.
- 44. Краскал WH, Уоллис WA. Использование рангов в однокритериальном дисперсионном анализе. Журнал Американской статистической ассоциации. 1952; 47: 583–621.
- 45. Спирмен К. Доказательство и измерение связи между двумя вещами. К. Спирмен, 1904 г.Am J Psychol. 1987; 100: 441–71. pmid: 3322052
- 46. Пирсон К. VII. Обратите внимание на регресс и наследование в случае двух родителей. Proc R Soc Lond. 1895; 58: 240–2.
- 47. Крамер Х. Математические методы статистики. Принстон: Издательство Принстонского университета; 1999.
- 48. Clooney AG, Fouhy F, Sleator RD, O ’Driscoll A, Stanton C, Cotter PD, et al. Сравнение яблок и апельсинов?: Секвенирование следующего поколения и его влияние на анализ микробиома.PLoS ONE. 2016; 11: e0148028. pmid: 26849217.
- 49. Kong HH. Важность деталей: разработка исследований микробиома кожи. J Invest Dermatol. 2016; 136: 900–2. pmid: 27107375.
- 50. Поллок Дж., Глендиннинг Л., Визедчанвет Т., Уотсон М. Безумие микробиома: попытка найти консенсус «Лучшая практика» для исследований микробиома 16S. Appl Environ Microbiol. 2018; 84. pmid: 29427429.
- 51. Коул Л.К. Анатомия и физиология собачьего уха. Vet Dermatol.2009; 20: 412–21. pmid: 20178478.
- 52. Уильям Х. Миллер Дж., Крейг Е. Гриффин Д.В., Кэмпбелл К.Л. Дерматология мелких животных Мюллера и Кирка. Elsevier Health Sciences; 2012.
- 53. Торрес С., Клейтон Дж. Б., Данцайсен Дж. Л., Уорд Т., Хуанг Х., Найтс Д. и др. На коже собак существуют разнообразные бактериальные сообщества, на которые влияет совместное проживание и время. PeerJ. 2017; 5: e3075. pmid: 28289569.
- 54. Клаэссон М.Дж., Ван К., О’Салливан О., Грин-Диниз Р., Коул Дж. Р., Росс Р. П. и др.Сравнение двух технологий секвенирования следующего поколения для определения очень сложного состава микробиоты с использованием тандемных вариабельных участков гена 16S рРНК. Nucleic Acids Res. 2010; 38: e200. pmid: 20880993.
- 55. Song SJ, Lauber C, Costello EK, Lozupone CA, Humphrey G, Berg-Lyons D, et al. Сожительствующие члены семьи разделяют микробиоту друг с другом и со своими собаками. Элиф. 2013; 2: e00458. pmid: 23599893.
- 56. Домингес-Белло М.Г., Костелло Е.К., Контрерас М., Магрис М., Идальго Г., Фирер Н. и др.Способ доставки формирует приобретение и структуру исходной микробиоты в различных средах обитания новорожденных. Proc Natl Acad Sci U S. A. 2010; 107: 11971–5. pmid: 20566857.
- 57. Грёнроос М., Параджули А., Лайтинен О.Н., Рослунд М.И., Вари Х.К., Хюти Х. и др. Кратковременный прямой контакт с почвой и растительным материалом приводит к немедленному увеличению разнообразия микробиоты кожи. Microbiologyopen. 2018: e00645. pmid: 29808965.
- 58. Санжар Ф., Уивер А.Дж., Пикок Т.Дж., Нгуен Дж.К., Бранденбург К.С., Леунг КП.Идентификация структуры и функции метагеномики, связанных с временными изменениями у крыс ( Rattus norvegicus ), микробиома кожи во время здоровья и кожного ожога. J Burn Care Res. 2020; 41: 347–58. pmid: 31665423.
- 59. Куско А., Белэнджер Дж. М., Гершони Л., Ислас-Трехо А., Леви К., Медрано Дж. Ф. и др. Индивидуальные особенности и факторы окружающей среды формируют микробиоту кожи здоровых собак. Микробиом. 2017; 5: 139. pmid: 235.
- 60. Janssen PH.Выявление доминирующих таксонов почвенных бактерий в библиотеках генов 16S рРНК и 16S рРНК. Appl Environ Microbiol. 2006; 72: 1719–28. pmid: 16517615.
- 61. Корбелик Дж., Сингх А., Руссо Дж., Виз Дж. С.. Характеристика отической бактериальной микробиоты у собак с наружным отитом по сравнению со здоровыми людьми. Vet Dermatol. 2019. pmid: 30828896.
- 62. Chermprapai S, Ederveen THA, Broere F, Broens EM, Schlotter YM, van Schalkwijk S, et al. Бактериальный и грибковый микробиом кожи здоровых собак и собак с атопическим дерматитом и влияние местной антимикробной терапии, предварительное исследование.Vet Microbiol. 2019; 229: 90–9. pmid: 30642603.
- 63. Mazhar S, Hill C, McAuliffe O. Род Macrococcus: взгляд на его биологию, эволюцию и взаимосвязь со стафилококком. Adv Appl Microbiol. 2018; 105: 1–50. pmid: 30342720.
- 64. Коттинг К., Штраус С., Родригес-Кампос С., Ростахер А., Фишер Н.М., Роосье П.Дж. и др. Macrococcus canis и M . caseolyticus у собак: встречаемость, генетическое разнообразие и устойчивость к антибиотикам.Vet Dermatol. 2017; 28: 559 – e133. pmid: 28748533.
- 65. Gobeli Brawand S, Cotting K, Gómez-Sanz E, Collaud A, Thomann A, Brodard I, et al. Macrococcus canis sp. nov., кожная бактерия, связанная с инфекциями у собак. Int J Syst Evol Microbiol. 2017; 67: 621–6. pmid: 276.
- 66. La Fuente R de, Suarez G, Ruiz Santa Quiteria JA, Meugnier H, Bes M, Freney J, et al. Идентификация коагулазонегативных стафилококков, выделенных от ягнят, как Staphylococcus caseolyticus .Comp Immunol Microbiol Infect Dis. 1992; 15: 47–52. pmid: 1547620
- 67. Гомес-Санс Э., Швенденер С., Томан А., Гобели Браванд С., Перретен В. Первая хромосома стафилококковой кассеты mec, содержащая генный комплекс, несущий mecB, независимый от транспозона Tn6045, в изоляте Macrococcus caseolyticus от собачьей инфекции. Антимикробные агенты Chemother. 2015; 59: 4577–83. pmid: 25987634.
- 68. Gonzales AJ, Bowman JW, Fici GJ, Zhang M, Mann DW, Mitton-Fry M.Оклацитиниб (APOQUEL®) – новый ингибитор киназы Януса, обладающий активностью против цитокинов, вызывающих аллергию. J Vet Pharmacol Ther. 2014; 37: 317–24. pmid: 24495176.
- 69. Видмер Дж., Феррер Л., Фаврот С., Папс Дж., Хьюн К., Оливри Т. Глюкокортикостероиды и циклоспорин существенно не влияют на микробиоту кожи собак. BMC Vet Res. 2018; 14:51. pmid: 29471815.
- 70. Yu G, Fadrosh D, Goedert JJ, Ravel J, Goldstein AM. Вложенные ошибки ПЦР в интерпретации структуры микробного сообщества в наборах данных последовательностей генов 16S рРНК.PLoS ONE. 2015; 10: e0132253. pmid: 26196512.
- 71. Tremblay J, Singh K, Fern A, Kirton ES, He S, Woyke T. и др. Влияние праймера и платформы на секвенирование метки 16S рРНК. Front Microbiol. 2015; 6: 771. pmid: 26300854.
- 72. Уолтерс В., Хайд Э. Р., Берг-Лайонс Д., Акерманн Г., Хамфри Г., Парада А. и др. Усовершенствованный бактериальный ген 16S рРНК (V4 и V4-5) и праймеры для генов-маркеров внутреннего транскрибированного спейсера грибов для исследований микробного сообщества. mSystems. 2016; 1.pmid: 27822518.
- 73. Wear EK, Wilbanks EG, Nelson CE, Carlson CA. Выбор праймера влияет на численность конкретной популяции, но не на динамику сообщества в ежемесячном анализе ампликона гена 16S рРНК во временном ряду прибрежного морского бактериопланктона. Environ Microbiol. 2018; 20: 2709–26. pmid: 29521439.
- 74. Капоне К.А., Дауд С.Е., Стаматас Г.Н., Николовски Дж. Разнообразие микробиома кожи человека в раннем возрасте. J Invest Dermatol. 2011; 131: 2026–32. pmid: 21697884.
- 75. Скотти Э., Буэ С., Ло Сассо Дж., Занетти Ф., Белкастро В., Пуссен С. и др. Изучение микробиома в условиях здоровья и болезней. Токсикологические исследования и применение. МикробиологияOpen. 2017; 1: 239784731774188.
- 76. О Дж., Конлан С., Полли Э. К., Сегре Дж. А., Конг Х. Х. Изменения в микробиоте кожи и носа у здоровых детей и взрослых. Genome Med. 2012; 4: 1–11. pmid: 22257447
- 77. Lehtimäki J, Sinkko H, Hielm-Björkman A, Salmela E, Tiira K, Laatikainen T. и др.Микробиота кожи и аллергические симптомы связаны с воздействием микробов окружающей среды. Proc Natl Acad Sci U S. A. 2018; 115: 4897–902. pmid: 29686089.
Праймер, конвейеры, параметры: вопросы секвенирования гена 16S рРНК
Абстрактные
Секвенирование гена 16S рРНК короткого ампликона в настоящее время является предпочтительным методом для исследований микробиомов. Однако сравнительные исследования различий в процедурах немногочисленны. Мы секвенировали образцы стула человека и имитировали сообщества с возрастающей сложностью, используя множество широко используемых протоколов.Короткие ампликоны, нацеленные на различные вариабельные области (V-области) или их диапазоны (V1-V2, V1-V3, V3-V4, V4, V4-V5, V6-V8 и V7-V9), были исследованы на предмет различий в исходе композиции. из-за выбора грунтовки. Далее, влияние кластеризации (операционные таксономические единицы [OTUs], OTU нулевого радиуса [zOTUs] и варианты последовательностей ампликонов [ASVs]), различных баз данных (GreenGenes, Ribosomal Database Project, Silva, геномная база данных 16S рРНК и The All-Species Living Tree), а также биоинформатические параметры по таксономическому назначению.Мы представляем систематическое сравнение всех обычно используемых V-областей с использованием хорошо зарекомендовавших себя праймеров. Хотя известно, что выбор праймера оказывает значительное влияние на полученный микробный состав, мы показываем, что микробные профили, полученные с использованием различных пар праймеров, нуждаются в независимой проверке эффективности. Кроме того, сравнение наборов данных по V-регионам с использованием разных баз данных может вводить в заблуждение из-за различий в номенклатуре (например, Enterorhabdus против Adlercreutzia) и различной точности классификации вплоть до уровня родов.В целом, определенные, но важные таксоны не подбираются определенными парами праймеров (например, Bacteroidetes пропущены при использовании праймеров 515F-944R) или из-за используемой базы данных (например, Acetatifactor в GreenGenes и базы данных 16S рРНК на основе генома). Мы обнаружили, что соответствующее усечение ампликонов имеет важное значение, и для каждого исследования необходимо тестировать различные комбинации усеченной длины. Наконец, настоятельно рекомендуются конкретные фиктивные сообщества достаточной и адекватной сложности. ВАЖНОСТЬ При секвенировании гена 16S рРНК было обнаружено, что определенные роды бактерий недостаточно представлены или даже отсутствуют в таксономических профилях при использовании неподходящих комбинаций праймеров, устаревших баз данных ссылок или неадекватных настроек конвейера.Что касается последнего, пороги качества, а также биоинформатические параметры (то есть подход кластеризации, конвейер анализа и конкретные настройки, такие как усечение) являются причиной ряда наблюдаемых различий между исследованиями. Выводы, сделанные путем сравнения одного набора данных с другим (например, между публикациями), кажутся проблематичными и требуют независимой перекрестной проверки с использованием совпадающих V-областей и единообразной обработки данных. Поэтому мы подчеркиваем важность продуманного дизайна исследования, включающего достаточно сложные имитирующие стандарты и соответствующий выбор V-области для интересующей выборки.Предварительно необходимо проверить использование конвейеров обработки и параметров.
Почему в новейших самолетах F-16 ВВС США не используются эти высокотехнологичные топливные баки?
Топливо – это вездесущий призрак, который нависает над каждым пилотом. Это здорово, когда тебе этого достаточно, и ужасно, когда нет. Это особенно актуально для тактических истребителей, которые, как известно, прожорливы. Конформные топливные баки (CFT) становятся все более популярным способом увеличения дальности полета существующих истребителей, таких как F-16, без существенного влияния на скорость и маневренность самолета.Многие зарубежные операторы используют CFT на блоках 50/52 и более поздних версиях Vipers, но почему американские F-16 последней модели не обладают такой же роскошью?
Конформные топливные баки– это не только последние модели F-16, F-15 давно использует собственные CFT, а CFT разрабатываются для Gripen, Super Hornet, Rafale, Ching Kuo и Eurofighter. Усовершенствованные версии МиГ-29 также были оснащены верхним конформным топливным баком, а китайский J-10 разрабатывает вариант CFT.
CFT F-16 весят всего 900 фунтов в комплекте и могут нести 450 галлонов, около 3050 фунтов реактивного топлива.Учитывая, что F-16 вмещает около 7000 фунтов внутреннего топлива, только CFT предлагают увеличение расхода топлива на 43,5 процента при небольшом дополнительном сопротивлении. Кроме того, они не занимают драгоценные станции хранения под крыльями или брюхом самолета, как традиционные резервуары.
G / O Media может получить комиссию
Пилоты довольно четко заявили о том, насколько мало CFT F-16 влияет на пресловутую динамику F-16. Майор Тимоти Макдональд, пилот проекта ВВС США по испытаниям F-16 CFT на авиабазе Эглин, описал добавление с болтовым креплением следующим образом:
«CFT практически не влияют на известные характеристики F-16.Комплект CFT несет на 50 процентов больше топлива, чем внешний топливный бак по средней линии, но имеет только 12 процентов сопротивления. Самолет сохраняет полную способность 9 g и диапазон полета с установленными CFT. Воздействие лобового сопротивления очень мало, менее одного процента в боевой конфигурации в крейсерских условиях ».
Стивен Бартер, пилот проекта Lockheed CFT во время их разработки, также заявил:
«Летные качества F-16 с CFT практически не изменились по сравнению с самолетом без CFT.Для большинства условий боевого полета создается впечатление, что CFT вообще не существует. Самый верный способ узнать, установлены ли CFT, – это посмотреть через плечо ».
F-16 CFT разработаны для полного диапазона полета реактивного самолета, до 9 g, максимального угла атаки, бокового скольжения и максимальной скорости крена, и они почти не влияют на маневренность или скорость ниже максимальной, где истребители тратят огромное количество времени. большую часть своего времени. Даже выше единицы, они имеют меньшее влияние на производительность и вмещают больше топлива, чем средний бак.
В то время как Израиль, Турция, Польша, Греция, Марокко, Пакистан, Оман, Сингапур и ОАЭ активно используют CFT на своих самолетах F-16, США полностью обошли этот вариант.
Принимая во внимание, что F-16 ВВС США никогда не летают в бою без нескольких внешних топливных баков, что сильно ухудшает характеристики самолета, разве CFT не является проблемой? Казалось бы, так, но ВВС США используют некоторую, казалось бы, отсталую логику, когда дело доходит до того, чтобы не закупать их для своего флота F-16 Block 50/52, который является самым современным и самым молодым субфлотом F-16 в мире. инвентарь ВВС США.
Судя по тому, что я собирал на протяжении многих лет по этой теме, разговаривая с пилотами напрямую и время от времени встречая упоминания о решении в публикациях, ВВС США оправдывают отказ от закупки CFT, потому что у них уже есть так много самолетов-заправщиков. Кроме того, в крупном воздушном сражении им не нужны дополнительные оружейные станции, освобожденные за счет включения CFT, как большое преимущество, потому что они могут просто назначить больше F-16 для набора целей. Для нанесения ударов на большие расстояния они могут использовать F-15E или даже бомбардировщик.
Эти обоснования, а также тот факт, что хотя CFT не препятствуют ежедневному линейному техническому обслуживанию и осмотрам при установке на самолет, их снятие для тяжелого обслуживания и фазовых осмотров занимает пару часов, кажется, все это является причиной отсутствия оборудования. Американские гадюки поздней модели с ЦФТ.
Теперь давайте подробнее рассмотрим это обоснование. Это правда, что у США головокружительное количество самолетов-заправщиков, всего около 450, из которых 59 являются удлинителями KC-10, а остальные – Stratotankers KC-135R.Эти самолеты также могут использоваться для грузовых операций, но в основном они обеспечивают дозаправку измученных жаждой военных самолетов США и их союзников. Между тем, F-16 по-прежнему представляет собой основу тактического боевого потенциала ВВС США: более 900 единиц все еще находятся в эксплуатации даже после того, как многие эскадрильи были закрыты из-за сокращения бюджета.
Более 250 из этих самолетов относятся к самолетам Block 50/52 F-16CJ, которые были поставлены оптимизированными для CFT. Другими словами, около 30 процентов самых современных и самых молодых F-16 ВВС США могут быть легко оснащены CFT, многие из которых обладают сложной системой подавления ПВО противника / уничтожения ПВО противника (SEAD / DEAD). миссия.Эта сложная миссия является критически важной возможностью поддержки, которая необходима для обеспечения безопасности самолетов ближнего и дальнего действия против врага даже с элементарной системой противовоздушной обороны.
F-16, с его ограниченной дальностью действия и «незавершенной» системой целеуказания (HTS), никогда не был идеально подходил для этой миссии, только действительно проявил себя в ней после ключевых обновлений авионики и вооружения за последнее время. десять лет или около того. Добавление еще примерно 40 процентов внутреннего топлива помогло бы сделать реактивный самолет лучшим Wild Weasel, каким он только мог быть, и дало бы ему все необходимое для работы зенитных систем, пока ударные и противовоздушные самолеты выполняли свои задачи в воздушное пространство противника.
Добавление почти 50 процентов внутреннего топлива к 25 процентам существующего парка F-16 ВВС США дает множество преимуществ как в тактическом, так и в стратегическом плане. Во-первых, значительно увеличено время ожидания и дальность действия. Это позволит этим F-16 не быть так тесно привязаны к воздушным заправщикам, как сегодня, и им придется отправляться примерно каждый час для дозаправки, находясь над районом боевых действий, что значительно усложняет планирование и может оставить брешь в ключевой области поле битвы. Это особенно важно при выполнении миссий критической поддержки с воздуха (CAS) и Wild Weasel.
В миссиях средней и малой дальности CFT означают большую маневренность, меньшее сопротивление лобовому сопротивлению и большее количество оружия, доступного на самолет, по сравнению с полетом на F-16 с громоздкими танками под крылом. В дальних миссиях, где подкрыльевые сбрасывающие баки соединены с CFT, это означает увеличенную дальность и время стоянки.
Что касается идеи о том, что огромные силы танкеров ВВС США делают CFT менее актуальными, это кажется очень близоруким взглядом на расширенные возможности CFT, и это несколько архаичный взгляд на текущие реалии воздушного боя.
Танкерный флот Америки со временем сократится. KC-135, многим из которых старше 50 лет, не могут летать вечно, и найти финансирование для их индивидуальной замены на KC-46 будет практически невозможно. Даже тяжеловесные удлинители KC-10 в последнее время не выходят за рамки бюджета просто из-за их стоимости эксплуатации.
Имейте в виду, что это происходит потому, что ВВС США планируют закупить более 1700 F-35A, каждый из которых имеет в два с половиной раза больше внутреннего топлива, чем их аналоги F-16.Это не было бы проблемой, если бы новые малозаметные самолеты имели удвоенную дальность или боевой радиус, но это не так. Таким образом, они будут сильно зависеть от танкерного топлива для выполнения своей миссии, как и их предшественники F-16, только они намного сильнее жаждут. Это окажет большее давление на сокращающийся танкерный флот во время будущих конфликтов.
Затем у вас есть предполагаемый «разворот Пентагона в сторону Тихого океана», где большие расстояния и реалии ограниченной логистики, аскетичного базирования и инфраструктуры поддержки – самые большие проблемы, которые необходимо преодолеть.Разрешить американским самолетам Block 50/52 F-16C / D действовать более независимо от танкеров на больших дальностях – логичный ход, если к этой стратегии будут относиться серьезно.
Кроме того, существует проблема уязвимости танкеров, поскольку они являются крупными целями, неспособными защитить себя, и которым поручено действовать в пределах нескольких сотен миль от своих коротконогих иждивенцев-истребителей. Во время войны с противником, обладающим способностью блокировать доступ и блокировать территорию, это может означать длинные «танкерные мосты», перебрасывающие истребители с удаленных баз на линию фронта.Это также означает, что сами танкеры будут развернуты далеко вперед. Таким образом, они будут уязвимы для вражеских атак, особенно во время операций с множеством врагов или тех, где используются передовые малозаметные вражеские самолеты. Таким образом, чем дальше эти танкеры могут быть отброшены за счет увеличения органической дальности действия наших боевых самолетов, тем в большей безопасности они будут во время такого конфликтного конфликта.
Также имейте в виду, что на обширных просторах Тихого океана, даже во время ограниченного конфликта между государствами, если противник убивает танкер, он также может убить всех истребителей, зависящих от этого танкера, поскольку им придется отвлекаться. на базы, уязвимые для вражеской атаки, или когда закончилось топливо и кончилось в океане
Наконец, у вас есть стоимость. Бензин для цистерн стоит от 25 до 35 долларов за галлон (некоторые утверждают, что это ближе к 50 долларам), если учесть расходы, связанные с дозаправкой в воздухе. Это по сравнению с примерно 5-6 долларами, когда самолет заправляется на земле. Во время длительных боевых действий низкой интенсивности или во время тренировок использование топлива наземного базирования в большей степени, чем танкеров, является гораздо более экономичным способом ведения боевых действий. Это также даст больше времени на вылет во время обучения, когда не используются средства дозаправки в воздухе.Это означает меньшее количество «поворотов» самолета и гораздо большую эффективность при выделении пилотам требуемых часов обучения. Меньше вылетов, но более длительные, также легче по технике.
Самое лучшее в CFT – это то, что они съемные. Для некоторых операций, если они будут сочтены ненужными, самолет можно оставить без них, а для других задач они могут быть установлены. Например, над Афганистаном, где настойчивость во время миссий CAS была ключевой, и не было угрозы воздух-воздух или повышенной опасности земля-воздух, транспортировка газа на 40% больше при возможности носить с собой больше оружия кажется довольно логичным преимуществом. .Тем не менее, даже во время боевых действий, где есть надежная тяга земля-воздух и воздух-воздух, любое небольшое препятствие, которое CFT создают для маневренности, которая, по-видимому, почти ничто, преодолевается за счет возможности дольше оставаться в бою. при более высоких настройках тяги.
Кроме того, мы живем во времена ракет класса «воздух-воздух» с высокой прямой наводкой и буксируемых ложных целей, когда грубая маневренность становится все меньше и меньше преимуществ в бою. Вместо этого первостепенное значение имеют дальность полета, настойчивость и способность максимально эффективно использовать силовую установку самолета, когда это необходимо, и столько, сколько необходимо.
Утверждение, что ВВС США могут просто выставить больше F-16, чтобы уничтожить определенное количество целей и выполнить определенное количество патрулей, кажется невероятно безрассудным. Тот факт, что активы доступны, не означает, что их использование необходимо. Зачем подвергать риску больше экипажей и материалов, чем это абсолютно необходимо во время войны? Особенно с учетом того, что потеря всего одного F-16 поставит под угрозу гораздо больше личного состава во время боевой поисково-спасательной операции.
Дело в том, что самые молодые американские F-16 будут служить еще десятилетия, скорее всего, взяв на себя многие роли «хлеба с маслом», для которых использование стелс-истребителя стоимостью более 100 миллионов долларов абсолютно бессмысленно.
Имея все это в виду, разве не разумно немного ослабить зависимость американских истребителей от бензина для танкеров и дать экипажам F-16C / D Block 50/52 большую настойчивость и дальность действия за счет добавления CFT? Особенно учитывая, что способность доказана, закреплена по своей природе, имеет небольшие неблагоприятные последствия, когда дело доходит до маневренности, и уже доступна в продаже?
Логика скажет «да», но ВВС США почему-то говорят «нет».
Свяжитесь с автором по телефону Tyler @ jalopnik.ком .
Фотографии: Турецкий F-16 wikicommons / SAC Helen Farrer RAF Mobile News Team, Сингапур F-16 Wikicommpns / Лухай Вонг, польский F-16 Wikicommons / SFJZ, верхний выстрел IAF / IDF, нижний выстрел через Lockheed Martin все остальные фото через USAF.
% PDF-1.4 % 214 0 объект > эндобдж xref 214 240 0000000016 00000 н. 0000006314 00000 н. 0000006399 00000 н. 0000006534 00000 н. 0000008208 00000 н. 0000008743 00000 н. 0000008780 00000 н. 0000008828 00000 н. 0000008876 00000 н. 0000008923 00000 н. 0000008971 00000 п. 0000009018 00000 н. 0000009066 00000 н. 0000009113 00000 п. 0000009161 00000 п. 0000009208 00000 н. 0000009256 00000 н. 0000009303 00000 п. 0000009351 00000 п. 0000009398 00000 п. 0000009446 00000 н. 0000009493 00000 п. 0000009541 00000 н. 0000009588 00000 н. 0000009636 00000 н. 0000009683 00000 н. 0000009731 00000 н. 0000009778 00000 п. 0000009826 00000 н. 0000009873 00000 н. 0000009921 00000 н. 0000009968 00000 н. 0000010016 00000 п. 0000010063 00000 п. 0000010111 00000 п. 0000010158 00000 п. 0000010206 00000 п. 0000010253 00000 п. 0000010301 00000 п. 0000010348 00000 п. 0000010395 00000 п. 0000010442 00000 п. 0000010489 00000 п. 0000010536 00000 п. 0000010584 00000 п. 0000010631 00000 п. 0000010679 00000 п. 0000010726 00000 п. 0000010774 00000 п. 0000010821 00000 п. 0000010868 00000 п. 0000010915 00000 п. 0000010962 00000 п. 0000011009 00000 п. 0000011056 00000 п. 0000011103 00000 п. 0000011150 00000 п. 0000011197 00000 п. 0000011244 00000 п. 0000011291 00000 п. 0000011338 00000 п. 0000011385 00000 п. 0000011432 00000 п. 0000011479 00000 п. 0000011526 00000 п. 0000011573 00000 п. 0000011620 00000 п. 0000011667 00000 п. 0000011714 00000 п. 0000011761 00000 п. 0000011808 00000 п. 0000011855 00000 п. 0000011902 00000 п. 0000011949 00000 п. 0000011996 00000 п. 0000012043 00000 п. 0000012090 00000 н. 0000012137 00000 п. 0000012184 00000 п. 0000012231 00000 п. 0000012278 00000 н. 0000012325 00000 п. 0000012372 00000 п. 0000012419 00000 п. 0000012466 00000 п. 0000012513 00000 п. 0000012560 00000 п. 0000012607 00000 п. 0000012654 00000 п. 0000012701 00000 п. 0000012748 00000 п. 0000012795 00000 п. 0000012842 00000 п. 0000012889 00000 п. 0000012936 00000 п. 0000012983 00000 п. 0000013030 00000 н. 0000013077 00000 п. 0000013124 00000 п. 0000013171 00000 п. 0000013218 00000 п. 0000013266 00000 п. 0000013313 00000 п. 0000013361 00000 п. 0000013408 00000 п. 0000013456 00000 п. 0000013503 00000 п. 0000013551 00000 п. 0000013598 00000 п. 0000013646 00000 п. 0000013693 00000 п. 0000013741 00000 п. 0000013788 00000 п. 0000013836 00000 п. 0000013883 00000 п. 0000013931 00000 п. 0000013978 00000 п. 0000014026 00000 п. 0000014073 00000 п. 0000014121 00000 п. 0000014350 00000 п. 0000014428 00000 п. 0000014998 00000 п. 0000017692 00000 п. 0000020371 00000 п. 0000098883 00000 п. 0000455793 00000 н. 0000456224 00000 н. 0000456655 00000 н. 0000456948 00000 н. 0000457418 00000 п. 0000457711 00000 п. 0000458184 00000 п. 0000458489 00000 н. 0000458977 00000 н. 0000459312 00000 н. 0000459755 00000 н. 0000460114 00000 п. 0000460530 00000 н. 0000460895 00000 н. 0000461320 00000 н. 0000461694 00000 н. 0000462119 00000 п. 0000462484 00000 н. 0000462897 00000 н. 0000463253 00000 н. 0000463699 00000 н. 0000464016 00000 н. 0000464435 00000 п. 0000464743 00000 н. 0000465174 00000 н. 0000465482 00000 н. 0000465904 00000 н. 0000466215 00000 н. 0000466628 00000 н. 0000466942 00000 н. 0000467349 00000 н. 0000467660 00000 н. 0000468061 00000 н. 0000468378 00000 н. 0000468755 00000 н. 0000469072 00000 н. 0000469458 00000 п. 0000469772 00000 н. 0000470167 00000 н. 0000470520 00000 н. 0000470915 00000 н. 0000471253 00000 н. 0000471648 00000 н. 0000472025 00000 н. 0000472384 00000 н. 0000472746 00000 н. 0000473099 00000 н. 0000473434 00000 н. 0000473778 00000 н. 0000474134 00000 н. 0000474475 00000 н. 0000474822 00000 н. 0000475166 00000 н. 0000475510 00000 н. 0000475839 00000 н. 0000476180 00000 н. 0000476512 00000 н. 0000476853 00000 н. 0000477191 00000 н. 0000477526 00000 н. 0000477858 00000 н. 0000478205 00000 н. 0000478555 00000 н. 0000478893 00000 н. 0000479234 00000 н. 0000479575 00000 н. 0000479916 00000 н. 0000480266 00000 н. 0000480619 00000 п. 0000480948 00000 н. 0000481289 00000 н. 0000481648 00000 н. 0000481986 00000 н. 0000482321 00000 н. 0000482653 00000 н. 0000482994 00000 н. 0000483338 00000 н. 0000483682 00000 н. 0000484005 00000 н. 0000484337 00000 н. 0000484663 00000 н. 0000484986 00000 н. 0000485288 00000 н. 0000485617 00000 н. 0000485949 00000 н. 0000486284 00000 н. 0000486622 00000 н. 0000486963 00000 н. 0000487268 00000 н. 0000487612 00000 н. 0000487956 00000 н. 0000488318 00000 н. 0000488686 00000 н. 0000489024 00000 н. 0000489368 00000 н. 0000489808 00000 н. 00004 00000 н. 00004 00000 п. 00004
- 00000 н. 0000491301 00000 н. 0000491630 00000 н. 0000492058 00000 н. 0000492375 00000 п. 0000492800 00000 н. 0000493102 00000 п. 0000493527 00000 н. 0000493814 00000 н. 0000494230 00000 н. 0000494532 00000 н. 0000494966 00000 н. 0000495277 00000 н. 0000495729 00000 н. 0000496031 00000 н. 0000005096 00000 н. трейлер ] >> startxref 0 %% EOF 453 0 объект > поток xU {L [U [B – E + cĹ.@ 7uDOHq [8G!] Ka ٖ 岱 a @ 1yqn3 ۖ, $ | S
Метабаркодирование ДНК улавливает тонкие различия в сообществах лесных жуков после нарушения
Вы здесьЦитата
Лю, М., Бейкер, С.К. и Берридж, С.П. и Джордан, Дж. Дж. И Кларк, Л., метабаркодирование ДНК фиксирует тонкие различия в сообществах лесных жуков после нарушения, Restoration Ecology, 28, (6) pp. 1475-1484. ISSN 1061-2971 (2020) [Реферируемая статья]
Заявление об авторских правах
Copyright 2020 Общество экологического восстановления
DOI: doi: 10.1111 / rec.13236
Абстрактные
Метабаркодирование ДНК– это новый подход к мониторингу биоразнообразия, но остается неопределенность в отношении его способности обнаруживать тонкие различия в составе сообщества беспозвоночных, сравнимые с теми, которые можно получить на основе традиционной морфологической идентификации. В этом исследовании методы, основанные на метабаркодировании ДНК и морфологии, сравнивались в качестве инструментов для изучения того, повлияла ли история лесозаготовок на сообщества жуков в влажных эвкалиптовых лесах Тасмании.Мы сравнили 12 незарегистрированных участков спелых лесов с 12 соседними участками лесовосстановления, которые были вырублены примерно 55 лет назад. Количество видов, идентифицированных на основе морфологии (173), было близко к количеству операционных таксономических единиц нулевого радиуса (ZOTU), идентифицированных с помощью ДНК-метабаркодирования субъединицы I цитохром с оксидазы (COI, 176) и 16S рибосомной РНК (16S, 156). маркеры. Незначительные, но существенные различия в видовом составе жуков между возобновляемыми и незащищенными спелыми лесами были обнаружены с помощью методов метабаркодирования ДНК на основе морфологии и COI, но не метабаркодирования ДНК 16S.Наши результаты подтверждают пригодность митохондриальных COI для изучения биоразнообразия беспозвоночных. Небольшая потеря сигнала по сравнению с подходом, основанным на морфологии, может быть решена путем разработки более полных эталонных баз данных ДНК. Подтверждая, что восстановление лесов в течение 48-58 лет не привело к полному восстановлению сообществ зрелых лесных жуков, мы предполагаем, что метабаркодирование ДНК можно использовать для мониторинга биоразнообразия и выявления тонких различий в составе сообществ.
Сведения о товаре
Тип элемента: Ссылка на статью Ключевые слова: мониторинг биоразнообразия, жесткокрылые, субъединица I цитохром-соксидазы (COI), восстановление лесов, высокопроизводительное секвенирование, метабаркодирование, метабаркодирование ДНК, жуки, лес, сохранение Отдел исследований: Биологические науки Исследовательская группа: Генетика Область исследований: Геномика Подразделение целей: Управление окружающей средой Группа целей: Наземные системы и управление Область цели: Биоразнообразие суши UTAS Автор: Лю, M (г-н Mingxin Liu) UTAS Автор: Baker, SC (Dr Sue Baker) UTAS Автор: Burridge, CP (Associate Pr офессор Кристофер Берридж) UTAS Автор: Jordan, GJ (профессор Грег Джордан) UTAS Автор: Clarke, L (Dr Laurence Clarke) Идентификационный код: 141920 Год публикации: 2020 Финансовая поддержка: Австралийский исследовательский совет (LP140100075) Web of Science ® раз процитировано: 1 Автор депонирования: Plant Science Депонировано: 2020-12-03 Последнее изменение: 2021-03-16 Загрузки: 0 Только сотрудники хранилища: страница управления предметами
проблем в секвенировании гена 16S рРНК – Университет Южной Дании
TY – JOUR
T1 – Праймер, конвейеры, параметры
T2 – Проблемы в секвенировании гена 16S рРНК
AU – Abellan-Schneyder, Isabel
МатчаAU , Моника С.
AU – Reitmeier, Sandra
AU – Sommer, Alina
AU – Sewald, Zeno
AU – Baumbach, январь
AU – List, Markus
AU – Neuhaus, Klaus
Copyright: Neuhaus, Klaus 9000 –
Авторские права © 2021 Абеллан-Шнейдер и др. Авторские права: Эта запись получена из MEDLINE / PubMed, базы данных Национальной медицинской библиотеки США
PY – 2021/2/24
Y1 – 2021/2/24
N2 – Секвенирование гена 16S рРНК короткого ампликона в настоящее время является метод выбора для исследований микробиомов.Однако сравнительные исследования различий в процедурах немногочисленны. Мы секвенировали образцы стула человека и имитировали сообщества с возрастающей сложностью, используя множество широко используемых протоколов. Короткие ампликоны, нацеленные на различные вариабельные области (V-области) или их диапазоны (V1-V2, V1-V3, V3-V4, V4, V4-V5, V6-V8 и V7-V9), были исследованы на предмет различий в исходе композиции. из-за выбора грунтовки. Далее, влияние кластеризации (операционные таксономические единицы [OTUs], OTU нулевого радиуса [zOTUs] и варианты последовательностей ампликонов [ASVs]), различных баз данных (GreenGenes, Ribosomal Database Project, Silva, геномная база данных 16S рРНК и The All-Species Living Tree), а также биоинформатические параметры по таксономическому назначению.Мы представляем систематическое сравнение всех обычно используемых V-областей с использованием хорошо зарекомендовавших себя праймеров. Хотя известно, что выбор праймера оказывает значительное влияние на полученный микробный состав, мы показываем, что микробные профили, полученные с использованием различных пар праймеров, нуждаются в независимой проверке эффективности. Кроме того, сравнение наборов данных по V-регионам с использованием разных баз данных может вводить в заблуждение из-за различий в номенклатуре (например, Enterorhabdus против Adlercreutzia) и различной точности классификации вплоть до уровня родов.В целом, определенные, но важные таксоны не подбираются определенными парами праймеров (например, Bacteroidetes пропущены при использовании праймеров 515F-944R) или из-за используемой базы данных (например, Acetatifactor в GreenGenes и базы данных 16S рРНК на основе генома). Мы обнаружили, что соответствующее усечение ампликонов имеет важное значение, и для каждого исследования необходимо тестировать различные комбинации усеченной длины. Наконец, настоятельно рекомендуются конкретные фиктивные сообщества достаточной и адекватной сложности. ВАЖНОСТЬ При секвенировании гена 16S рРНК было обнаружено, что определенные роды бактерий недостаточно представлены или даже отсутствуют в таксономических профилях при использовании неподходящих комбинаций праймеров, устаревших баз данных ссылок или неадекватных настроек конвейера.Что касается последнего, пороги качества, а также биоинформатические параметры (то есть подход кластеризации, конвейер анализа и конкретные настройки, такие как усечение) являются причиной ряда наблюдаемых различий между исследованиями. Выводы, сделанные путем сравнения одного набора данных с другим (например, между публикациями), кажутся проблематичными и требуют независимой перекрестной проверки с использованием совпадающих V-областей и единообразной обработки данных. Поэтому мы подчеркиваем важность продуманного дизайна исследования, включающего достаточно сложные имитирующие стандарты и соответствующий выбор V-области для интересующей выборки.Предварительно необходимо проверить использование конвейеров обработки и параметров.
AB – Секвенирование гена 16S рРНК короткого ампликона в настоящее время является методом выбора для исследований микробиомов. Однако сравнительные исследования различий в процедурах немногочисленны. Мы секвенировали образцы стула человека и имитировали сообщества с возрастающей сложностью, используя множество широко используемых протоколов. Короткие ампликоны, нацеленные на различные вариабельные области (V-области) или их диапазоны (V1-V2, V1-V3, V3-V4, V4, V4-V5, V6-V8 и V7-V9), были исследованы на предмет различий в исходе композиции. из-за выбора грунтовки.Далее, влияние кластеризации (операционные таксономические единицы [OTUs], OTU нулевого радиуса [zOTUs] и варианты последовательностей ампликонов [ASVs]), различных баз данных (GreenGenes, Ribosomal Database Project, Silva, геномная база данных 16S рРНК и The All-Species Living Tree), а также биоинформатические параметры по таксономическому назначению. Мы представляем систематическое сравнение всех обычно используемых V-областей с использованием хорошо зарекомендовавших себя праймеров. Хотя известно, что выбор праймера оказывает значительное влияние на полученный микробный состав, мы показываем, что микробные профили, полученные с использованием различных пар праймеров, нуждаются в независимой проверке эффективности.Кроме того, сравнение наборов данных по V-регионам с использованием разных баз данных может вводить в заблуждение из-за различий в номенклатуре (например, Enterorhabdus против Adlercreutzia) и различной точности классификации вплоть до уровня родов. В целом, определенные, но важные таксоны не подбираются определенными парами праймеров (например, Bacteroidetes пропущены при использовании праймеров 515F-944R) или из-за используемой базы данных (например, Acetatifactor в GreenGenes и базы данных 16S рРНК на основе генома). Мы обнаружили, что соответствующее усечение ампликонов имеет важное значение, и для каждого исследования необходимо тестировать различные комбинации усеченной длины.Наконец, настоятельно рекомендуются конкретные фиктивные сообщества достаточной и адекватной сложности. ВАЖНОСТЬ При секвенировании гена 16S рРНК было обнаружено, что определенные роды бактерий недостаточно представлены или даже отсутствуют в таксономических профилях при использовании неподходящих комбинаций праймеров, устаревших баз данных ссылок или неадекватных настроек конвейера. Что касается последнего, пороги качества, а также биоинформатические параметры (то есть подход кластеризации, конвейер анализа и конкретные настройки, такие как усечение) являются причиной ряда наблюдаемых различий между исследованиями.Выводы, сделанные путем сравнения одного набора данных с другим (например, между публикациями), кажутся проблематичными и требуют независимой перекрестной проверки с использованием совпадающих V-областей и единообразной обработки данных. Поэтому мы подчеркиваем важность продуманного дизайна исследования, включающего достаточно сложные имитирующие стандарты и соответствующий выбор V-области для интересующей выборки.